膜たんぱく質の判断機能は腸の判断機能へと受け継がれている。
腸の「判断機能」の起源を追求する中で、細胞膜=膜たんぱく質が重要な器官であること、その選択透過性が判断機能の起源になっているのではないかという部分まで見えてきました。
今回は、膜たんぱく質の選択透過性とその仕組みについて追求していきます。
細胞は、脂質二重層構造を持った細胞膜で内外が隔てられています。この性質だけでは、ホルモンやたんぱく質などの物質を取り込むことができません。
そこで、膜タンパク質の判断機能を活かして細胞膜を透過する膜輸送の仕組みをもっているのです。
では、具体的にどのように判断して取り込んでいるのでしょうか?
実は、小さな物質と大きな物質を取り込むのとで、その方法が異なります。
小さな物質の代表としてナトリウム-カリウムポンプ等のイオンポンプが該当します。
それらを取り込む場合は、これら専用の鍵穴(受容体)に、鍵(ナトリウムイオンやカリウムイオン)
が結合した際に輸送たんぱく質に変容して通過していきます。
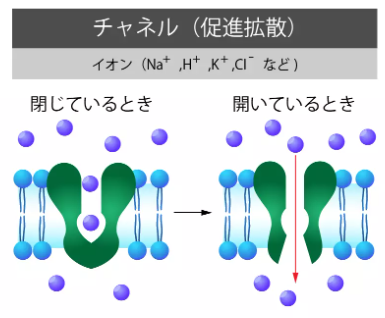
受験のミカタ(https://juken-mikata.net/how-to/biology/cell-membrane.html)様より転載
大きな物質の代表としてはホルモンやたんぱく質などがあります。
それらを取り込む場合は、細胞膜上の鍵穴(受容体)にホルモンやたんぱく質などの鍵(細胞外物質)が結合して初めて取り込みが行われます。
具体的には細胞膜がタンパク質等の大きな細胞外物質を取り込む形で陥入し、陥入した部分が細胞外物質を包みこむ形で細胞膜から分離して、細胞内に物質を取り込んでいます。
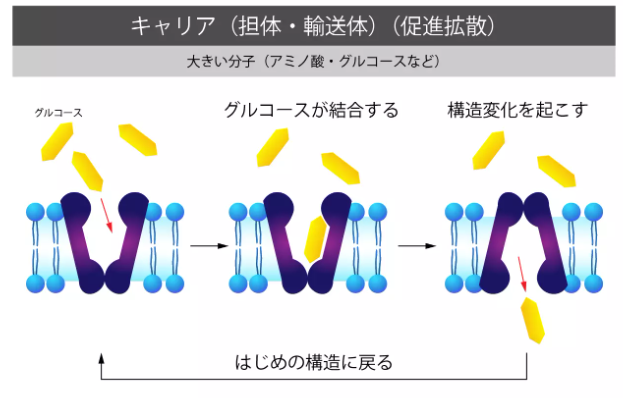
受験のミカタ(https://juken-mikata.net/how-to/biology/cell-membrane.html)様より転載
細胞は常に周囲からの様々なシグナル(細胞外物質)にさらされており、細胞の特性に従って、細胞膜に組み込まれた受容体(鍵穴)を変容させて選択的に応答しています。このようなシグナルの無限的組み合わせと、それにフィットする選択的応答方法(=受容体)が実現されています。
これら細胞内への取り込みこそ、膜たんぱく質が選択性を持って行われている、つまり判断していることを意味しています。
この膜輸送システムは、単細胞時代から存在するものです。
多細胞生物になった段階においても、それぞれの組織機能に応じた機能特化が行われていますが、基本的な構造は単細胞時代から変わっていないのです。
腸のもつ判断機能が膜たんぱく質の判断機能に由来しているという事が分かりますね。
続きを読む "膜たんぱく質の判断機能は腸の判断機能へと受け継がれている。"

■微生物と次代を生き抜く③~肥満の原因は腸内細菌が大きく関係している~
- 肥満はここ数十年における人類史上の大きな変化
現代はかつてなかった自己免疫疾患や、消化器系の不全、心の病、アレルギーが増えています。(http://www.seibutsushi.net/blog/)それに伴い目立つようになってきたものが、肥満です。
1950年代に、アメリカで突如体重増加が顕在化し、現在では、成人の1/3が過体重、1/9が肥満となっている。ヨーロッパでも、過去50年において平均して20%も体重が増えており、肥満はここ数十年における人類史上の大きな変化となっています。肥満の増加に伴って、糖尿病や心臓病も増加しています。

- 肥満の原因は腸内細菌が大きく関係している!
肥満の主な原因は、一般的には食べ過ぎと運動不足と言われているが、最近の研究によると、腸内細菌バランスが肥満と大きく関係してることが分かってきました。肥満の人と痩せている人の腸内環境を比べると、ある特定の腸内細菌のバランスが大きく異なる事が分かってきました。

- 吸収を抑えてくれる腸内細菌
もう一つは「アッカーマンシア・ムシニフィラ」という細菌。痩せた人では腸内微生物全体の4%を占めるが、太った人にはほとんどいないことが分かった。アッカーマンシア・ムシニフィラは、腸壁を覆う厚い粘膜層の表面にいる細菌で、腸壁細胞に働きかけて粘液の分泌を促している。この細菌が少ないと、粘液の層が薄くなり、リポ多糖が血液に入り込みやすくなり、肥満につながってしまう。つまり、アッカーマンシア・ムシニフィラは吸収を抑えてくれます。
- 食欲を抑えてくれる腸内細菌
まず一つは、「フィーカリバクテリウム・プラウスニッツィ」という細菌。日本人で肥満の人と痩せている人、10人ずつの腸内細菌を検査したところ、やせている人のフィーカリバクテリウム・プラウスニッツィの割合の平均は5.9%に対して、肥満の人は3.9%と少ないという結果が報告されている。この腸内細菌は「短鎖脂肪酸」をつくり、腸からGLP-1などさまざまなホルモンの分泌を促進することで、胃腸の動きをゆるやかにします。食べたものが胃腸をゆっくり通過すると、満腹感を得やすくなって食事量が減り、肥満が抑制されるのです。つまりフィーカリバクテリウム・プラウスニッツィは「もっと食べたい!」という欲を抑えてくれるのです。
- 腸内細菌がつくる短鎖脂肪酸を増やす「食物繊維」や「多糖」
ただし、肥満の人でも、腸内細菌に短鎖脂肪酸を多くつくらせることが可能です。 その方法の1つが、食物繊維を多く摂取することです。食物繊維は、腸内細菌によって短鎖脂肪酸につくり変えられるためです。食物繊維は、根菜類、豆類、きのこ類、海藻類などに多く含まれています。
また、最近の研究では、イモ類や寒天などに含まれる多糖という物質の一種が、腸内細菌によって、より短鎖脂肪酸につくり変えられやすいこともわかってきました。
肥満にならないために食べる量を抑える傾向がありますが、腸内細菌の働きや仕組みを知り、それを活かした食生活を整えることが健康への第一歩ですね!

続きを読む "■微生物と次代を生き抜く③~肥満の原因は腸内細菌が大きく関係している~"
生き物とは何か?~生き物の細胞分裂の仕組み~
今回は、生き物とはそもそも何なのか?を考えていきたいと思います。
生き物とは何か?という根源的な問いに答える前に、「サビ」と「カビ」の違いを押さえることで、生き物の共通項を導くことができます。


どちらもどんどん増殖していく様を想像すれば、同じような現象だと思われるかもしれませんが、まったく異なる現象です。少し考えてみましょう。
・サビは鉄に酸素が結びついた化学変化?しかし、カビも化学変化で増殖している。
・サビは無機質な感じがして、カビは生き物っぽい気がする。
・カビは自ら増えている。サビは自分では増えることができない。水にさらされるとか、環境の変化で増殖していく。
どうやら、「自ら分裂し、複製できること」が生き物とそうでないものの違いになりそうです。
しかし、そもそも分裂して複製するってどういうことなのでしょうか?
私たちの体の中でも起こっている現象をクローズアップしていきましょう。
続きを読む "生き物とは何か?~生き物の細胞分裂の仕組み~"
『外圧判断機能』の起源~細胞膜の仕組み/膜たんぱく質がポイント?~
「外圧判断機能」の起源を追求する中で、腸の起源から海綿動物へ、その体を構成する細胞の一つの襟細胞、さらに襟細胞は単細胞生物の中の立襟鞭毛虫と類似するとろこまで見えてきました。さて、立襟鞭毛虫から、さらに遡って「判断機能」を軸にさらに探ってみようと思います。
続きを読む "『外圧判断機能』の起源~細胞膜の仕組み/膜たんぱく質がポイント?~"
■微生物と次代を生き抜く(2)~腸内細菌の環境は生まれた時から決まってしまう?!~
●我々は腸内細菌と共生している
現代はかつてなかった自己免疫疾患や、消化器系の不全、心の病、肥満、アレルギーが増えています。(http://www.seibutsushi.net/blog/)
それには、腸内細菌が大きく関係しているようです。 人体には、500種類を超える細菌が存在し、その細胞の数は100兆個以上!!
人体を構成する細胞の数は37兆個程度と言われているので、細菌数は3倍以上! 体内に棲む細菌のうち、約9割が腸内に棲みついており、重さは約1キロ~2キロにもなります。 この腸内細菌が、免疫機能と深く関わっています。 人体に共生している細菌集団が人体の免疫システムと緊密に連絡して体を守ってくれているというのが最近解明されつつあり、人間を構成する存在として切っても切れないものになっているようです。
●腸内細菌はどこからやってくるの?
そもそもお母さんのお腹の中にいる赤ちゃんの腸内に細菌は棲んでいません。では、どこからやってくるのでしょうか。長い間、感染ルートは不明だったが、ヤクルト研究所によって解明が進んでいます。
【赤ちゃんからお母さんと同じビフィズス菌が見つかった】
ビフィズス菌の起源を探るために、最初に出産前の妊婦の糞便を採取し、次に出産した新生児の糞便を採取し、それぞれからビフィズス菌を取り出しました。双方のビフィズス菌の遺伝子を解読して、同じ菌なのか違う菌なのかを検証。
母親と新生児の糞便からビフィズス菌が多く取り出しされた自然分娩8組の母子について調べたところ、6組の母子について、母子間で同一のビフィズス菌の存在が明らかになりました。
個人差はあるものの、出生1週間以内に同一のビフィズス菌が検出された例が多くみられました。さらに、母親から受け継いだ菌は、生後3か月を過ぎても乳児の腸内に定着していて、伝播した菌が長期間にわたって乳児の腸管に生息していることも示されています。
次に、分娩様式の違いによってビフィズス菌の伝播が異なるかどうかに関して検証。
自然分娩12組と帝王切開5組の合計17組の母子の糞便を解析したところ、自然分娩で生まれた新生児の11人から、母親と同一のビフィズス菌を持っていることが確認されました。
母親から伝播した菌が消化官に定着したものと考えられます。
【産道を通る際にビフィズス菌が口から腸へ入っていく】
一方、帝王切開で生まれた5人の新生児からは、母親と同一のビフィズス菌は検出されず、母子伝播は観察されませんでした。また、新生児の糞便からビフィズス菌が検出されるまでの時期も遅く、1ヶ月を過ぎる頃になると菌数は少ないながらビフィズス菌が定着し始めました。
これらの違いは以下のように考えます。
自然分娩の際、母親のビフィズス菌が新生児の腸管に速やかに定着し、腸内で増殖することで、ビフィズス菌が優勢な腸内フローラを形成することができます。
一方、帝王切開では、分娩時に母親のビフィズス菌と出会うことがないため、母親のビフィズス菌を受け取ることができません。そのため新生児の腸内フローラにビフィズス菌が定着する時期が遅くなり、ビフィズス菌も母親由来のものではないと考えらました。
自然分娩と帝王切開の大きな違いといえば、産道を経由するか否かです。この時の研究では妊婦の膣の細菌を調べていないため、以下は推測に過ぎませんが、一般に膣内からビフィズス菌が検出されることから、母親の腸内ビフィズス菌が膣に伝播。産道を通過する際に、ビフィズス菌が胎児の口から腸内へと到達したと考えられます。

【帝王切開で生まれた新生児のビフィズス菌はどこから?】
では、帝王切開で生まれた新生児のビフィズス菌の由来は、どこからなのでしょう。
病院などの医療現場で、細菌が患者から患者へと水平伝播する事例は良く知られています。 そこで同じ病院で、帝王切開によって生まれた新生児の糞便を調べましたが、同一のビフィズス菌は得られず、医療従事者や病院の環境由来のビフィズス菌が水平伝播する事例はみられませんでした。
しかし、帝王切開で生まれた新生児も、生後1ヶ月程で定着がみられることから、口等の経路でビフィズス菌が体内に取り込まれていると推測されます。
●母乳には新生児の腸内環境を育む力がある
新生児が健やかに成長するには、良好な腸内環境を育んでいく必要があります。
その重要な役割を果たすのが母乳です。母乳にはビフィズス菌を増やす効果がある「ガラクトオリゴ糖」が含まれています。
さらに、病原体やウイルスの侵入を防ぐために深く関わっているIgA抗体も含まれています。
生まれたばかりの赤ちゃんは、自分でIgAを作ることができません。しかし、母乳によって赤ちゃんの体内にIgAを移行できるため、細菌やウイルスから身体を守ることができるのです!
他の腸内細菌も同様に伝播すると考えられ、定着は3歳までにほぼ決まると云われています。
現代の医療の世界や、母親の腸内環境の弱体化故か、帝王切開が増えているという現状より、生まれた時から腸内環境が決まってしまうとも云えるのではないでしょうか。
従って、母親は妊婦の段階から良好な腸内環境を維持することが大切であり、人間の健康の第一歩となります。
では、その腸はどんな役割を果たしているのでしょうか?
続きを読む "■微生物と次代を生き抜く(2)~腸内細菌の環境は生まれた時から決まってしまう?!~"
脂肪由来幹細胞(ASC)って何?
前回の記事http://www.seibutsushi.net/blog/2023/02/8607.htmlでは、昆虫がさなぎになるときにドロドロの状態になっており、その正体は何なのか?を追求していきました。ドロドロの状態は「幹細胞に戻っている」こと、加えてその元は「血液」ではないか?と仮説を立てました。
今回はその「幹細胞」について深めていきたいと思います。
最近の研究ではこの幹細胞は、骨髄ではなく、脂肪内に大量に潜んでいるという事実が明らかになってきています。幹細胞が何かカギを握っていそうです。
今回は「幹細胞」についてみていきたいと思います。
腸の起源?〜海綿動物の襟細胞と立襟鞭毛虫〜
多細胞の始まりは腸から?腸の起源とは?
これまで睡眠について追求していく中で睡眠にはメラトニンやセロトニンといったようなホルモンが大きく睡眠の質と関連しているということがわかってきました。ホルモンの分泌のバランスを整えることで質の良い睡眠を取ることが可能になってきます。
では、それの大本であるホルモンはどこからできるのでしょうか。
答えは「脳」ではなく「腸」からです。
腸は消化器官の一部という認識が一般的ですが、体の中でたくさんの役割を担っています。
腸の役割とは何でしょうか?
一つは前述した「食物の分解、吸収、運搬」です。多くの動物が食物から栄養を体内に吸収できるのは腸があるからですね。
腸には消化以外にも、もう一つ大きな役割があります。
それは上記のホルモン分泌のように腸自体が状況をつかみ、判断し、指令を出す「判断機能」です。
これまでは「脳」がすべてを判断し、処理していると考えられてきましたが、腸も外圧をつかみ、判断し、迷走神経を使い脳とすり合わせを行っているということが近年の研究で明らかになってきています。
他にも生物によっては腸で呼吸ができたりと、腸は体の中で本当に様々な役割を担っています。脳の追求をしていく中では切っても切り離せない「腸」。
今回は腸の起源にまでさかのぼり、腸の謎を紐解いていきます。
微生物と次代を生き抜く① ~プロローグ 病を抱えた現代人
人類の平均寿命は、ここ100年程の間に大きく変化しています。日本では、1900年の男女の平均寿命は44歳、現在では80歳を超えています。先進国を中心に各国とも同じような傾向にあります。

100年程前の人類の代表的な死因は、肺炎、結核、感染性下痢症でした。この100年程の間に、予防接種の浸透。医療現場の衛生概念の進化による医療現場での感染の減少。水道をはじめとする、公衆衛生の充実。抗生物質の利用などにより、感染による病の克服が著しく進みました。
その結果、乳幼児の死亡率は減少し、平均寿命も著しく伸長してきました。現在では、代表的な死因も、かつての細菌に感染して起きる病から、心臓病、癌、脳卒中へと変化しています。
我々の寿命は延びましたが、21世紀に入ると、先進国を皮切りに、アレルギー、自己免疫疾患、消化器系の不全、心の病、肥満が激増してきました。高齢化によって体が弱った人にこうした病が増えているのかと思いきや、そうではないのです。むしろ若い人々にこそ増えているのです。
身近でもこうした症状を抱える人は珍しくなく、かなりの割合の人がこれら慢性的な不調に陥っている事が、一見当たり前の様になっています。すぐ死に至るものではなくとも、理由がよくわからないままに心身の不調に陥り、長期に渡って苦しむ。死に至る症状に繋がる遠因にもなる。など、急増している現代病によって、我々は大なり小なり常時不調をきたしているのです。これらはほんの数十年前は稀な事象であり、ごく短期間の内に進行している、人類にとって異常な事態なのです。
これら21世紀以降急増している現代病は何故ここまで増えてきたのでしょうか?
続きを読む "微生物と次代を生き抜く① ~プロローグ 病を抱えた現代人"
古細菌(アーキア)はなぜ多様に進化することができたのか?
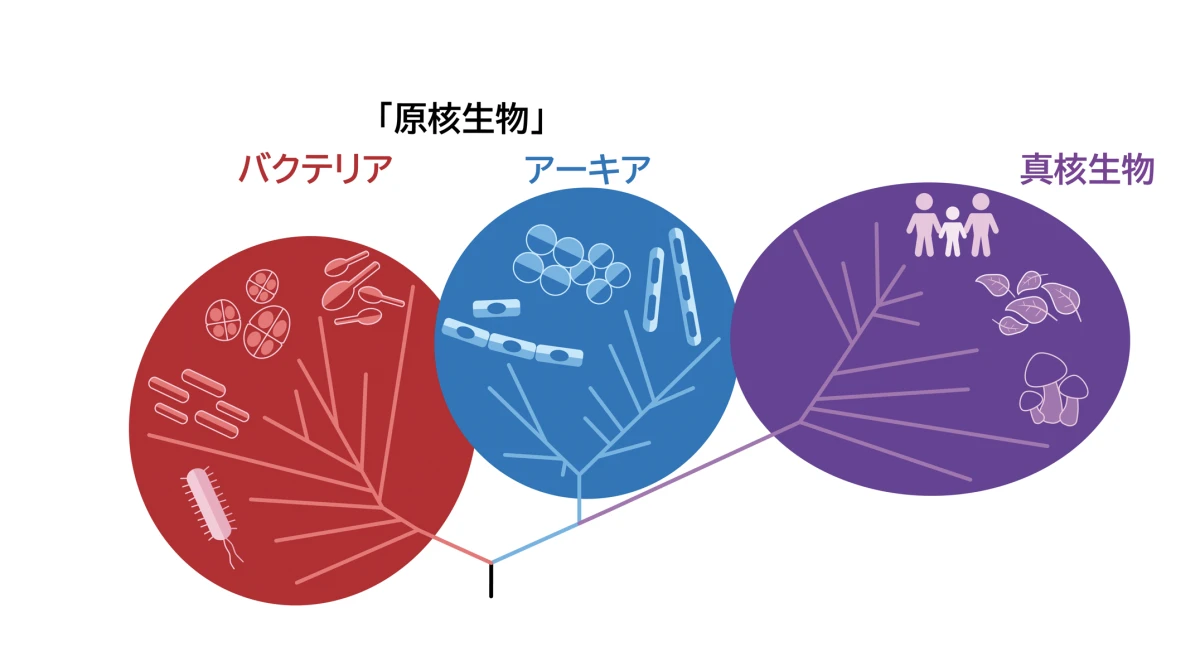
人類はもともと細菌だったということは現在ではよく知られていることです。細菌には、古細菌(以下、アーキア)と真正細菌(以下、バクテリア)の2系統が存在しています。しかし、哺乳類まで進化できたのはアーキアだと言われています。アーキアはなぜ多様に進化することができたのでしょうか。アーキアとバクテリアの違いから考えてみましょう。
40~35億年前、この時期に誕生したのが、アーキア、バクテリアの2系統の原核単細胞です。もともと地球には酸素がなかったので、両者とも0.5~2μm位のサイズの球状、棒状、または不定形の非常に小さな生物です。アーキアには、好熱菌、好塩菌、メタン生成金などがあります。バクテリアには、大腸菌、シアノバクテリア、枯草菌などがあります。これらの原核単細胞生物は、まだ地球に酸素がない時代に、硫黄や窒素を代謝してエネルギーを生み出していました。実は、人類も極めて少量ではありますが、硫黄を代謝しています。
アーキアとバクテリアの共通点は、DNAはむき出しの状態で核を持たないということです。しかし、アーキアは植物、哺乳類など多様に進化した真核生物へと分岐しましたが、バクテリアは微生物のままです。この違いは何なのでしょうか。
続きを読む "古細菌(アーキア)はなぜ多様に進化することができたのか?"
現代病:アルツハイマー型認知症の本当の原因に迫る②~神経を破壊するカルパイン~
現代病:アルツハイマー型認知症の本当の原因に迫る①では、アミロイドβの増加・蓄積を発生させるものとして、近年増加する人工物質の影響があることを明らかにしました。
しかし、アミロイドβの増加・蓄積だけであれば老人斑として誰でも起こる状況とも言えます。アルツハイマーの特徴でもある、徘徊・妄想・健忘など、明らかに通常の高齢化とは異なる症状は、何によって引き起こされているのでしょうか。

●カルパインが引き起こす、2つの事象
人工物質の増加により、アミロイドβが増加すると、具体的に脳内では、どのようなことが起こっているのでしょうか。
増加したアミロイドβは、通常、ネプライシンによって分解され、脳内から排出されていきます。しかし、人工物質等によって、アミロイドβが異常増加すると、ネプライシンを破壊する物質が増えてしまいます。それが『カルパイン』です。アミロイドβ増加により増えた脳内カルシウムを餌に、カルパインは過剰活性化していきます。
カルパインの過剰活性化により、引き起こされるのが『アミロイドβの蓄積』と『神経損傷の促進』です。