好熱古細菌の外圧認識⇒転写制御機構
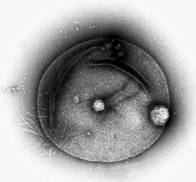
※Pyrococcus horikoshii OT3
今週は、GADV仮説~マリグラヌール~生体膜~エネルギー代謝・・・と、生命の起源に迫るエントリーが続いていますので、今日は認識機構の観点から考えてみたいと思います。
生命が「外の環境を認識して適応反応する」この奥深い仕組みはどのように形成されてきたのか?
気になる続きはポチっと押してからどうぞ 

![]()

まずは、パイロコッカス菌の研究事例を紹介します。
沖縄の深海底、100度以上の熱水噴出孔の近くに生息している好熱古細菌です。
●古細菌であるパイロコッカス菌が、栄養源(アミノ酸)の濃度を感知し、饗宴-飢餓モードに対応しながら遺伝子の転写を制御する「FFRP」を持つことを発見
※科学技術振興機構(JST)「地球上で最初に誕生した生命(コモノート)」の姿に一歩迫る発見(コモノートの代謝制御機構から生命の起源解明に向けて)
これまで本研究グループは古細菌のゲノム配列を決定し、また、これを解析する中で、各古細菌が数個から十数個の特定タイプの転写制御(注4)に関わるたんぱく質・FFRP(Feast/Famine Regulatory Protein:饗宴-飢餓・制御たんぱく質)を持つことを明らかにし、その構造や代謝制御機構解明に取り組んできました。
この結果、沖縄海溝の熱水チムニーに生息する古細菌・パイロコッカス菌のFFRPのひとつが、アミノ酸の一種である「リジン」の濃度で外界の栄養状態を感知し、これをバロメーターにして饗宴モードと飢餓モードに対応して会合状態を変化させること、そして、これに基づき代謝に関係する遺伝子群(全遺伝子の約20%)を制御する機構を持つことを明らかにしました。
パイロコッカス菌は、頭上の海から降り注ぐ魚等の残骸に含まれるたんぱく質から得たリジンなどのアミノ酸類を栄養源として生育し、栄養源が多いと増殖します。リジンと相互作用するとFFRPは図1のように8量体を形成(2量体4つが会合)し、FFRP遺伝子上流に位置するDNAに選択的に結合します。この結果、FFRP遺伝子の転写が抑制されて細胞中のFFRPの数が減少し、代謝関連遺伝子上流には結合できなくなるため、パイロコッカス菌の全代謝系は活性化されて”饗宴モード”に入り、菌は活発に増殖しはじめます。
一方、リジンが少ない低栄養状態では、FFRP8量体は2量体へと解離し、FFRP遺伝子上流部に位置するDNAとの結合は弱まります。この結果FFRP2量体の数は細胞あたり6000程度まで増加し、200ヵ所以上の遺伝子上流に結合して、これらの転写を抑制(図2)するため、代謝は”飢餓モード”に突入し、増殖が抑えられます。この代謝系の転写制御機構は、FFRPの構造や機能をさまざまな角度から研究したことにより解明されました。
よくできた仕組みですね ![]()
「FFRP」と呼ばれるタンパク質群が、アミノ酸(リジン)濃度を媒介として外的環境の変化を認識し、遺伝子の転写を制御しているわけです。
生命の外圧適応の観点からは、こうした「制御因子」(と受容体)が認識機構の要、DNA上の多くの遺伝子発現はその制御下にあると言えます。
もうひとつ注目点は、FFRPが真正細菌の大腸菌にも存在することです。
このことから、古細菌と真正細菌が分かれる前の共通祖先の段階から、こうした認識機構が存在していたことが推察されます。
つまり「外的環境(外圧)認識⇒適応反応」の最も原初的な回路である可能性が高い。
生命の誕生前夜=物質から生命へのあゆみが、「アミノ酸の重合」(マリソール→マリグラヌール様の物質) あたりからはじまったのだとすれば、アミノ酸の重合するメカニズムがベースとなって、(FFRPのような)アミノ酸濃度に応じたタンパク質変成のメカニズムを生み出し、それが外圧認識機構のベースとなっていったという仮説も考えられそうです。
(※生物の認識機構を考える上では、上記「外圧認識」ともうひとつ「同類認識」が重要になります。同類認識の起源も探っていくと興味深いテーマになりそうです。→参照記事:原始生命と群れ【仮説】 単細胞生物の同類認識)

トラックバック
このエントリーのトラックバックURL:
http://www.seibutsushi.net/blog/2009/01/660.html/trackback