2019-02-26
脳回路の仕組み11 構造認識と大脳連合野
・小脳や大脳の回路の全ては、瞬間瞬間の外識発で形成された専用回路である。
・しかし、瞬間瞬間の外識発で専用回路を形成してゆけば、忽ち脳容量の限界に達する。(脳容量に限界があるからこそ、使われない回路は新しい外識発の専用回路に乗っ取られ、塗り変えられてゆく)
・与えられた容量限界の中でより速く正体を掴むために生み出されたのが構造認識(共通項を抽出した類型認識や抽象概念や法則や原理)であり、そのために形成されていったのが大脳連合野である。
実際、大脳連合野は、同類情報が飛躍的に増大した共認動物において、更に共認機能を進化させて観念機能を生み出した観念動物において、著しく発達した。
・様々な認識群を構造認識で統合すれば、「どうする?何?何で?」の判断スピードが飛躍的に上昇する。更には、その体系化された認識群を使って新たな可能性(新認識)を発掘する地平が開かれる。かくして、全ての認識は構造認識へと先端収束してゆくことになる。
・共認動物→観念動物は、生存上の極限状態に追い込まれて本能を超えた共認機能や、その共認機能を超えた観念機能を形成する過程において、どうする?⇒何?なんで?の探求機能を飛躍的に発達させたが、この探求を主要に担ったのが大脳連合野である。
・そこでは、どうする?⇒何?なんで?の探求に関連する情報を突き合わせて統合する必要がある。その突き合わせのために形成されたのが、5秒~10秒だけ情報を再現する(そして突き合わせる)作業記憶の回路である(その機能は反復体(海馬)に似ているが、反復体よりも遥かに短時間で消去されてゆく)。
・そして、部分的に統合された認識(初歩的な構造認識)とその関連情報の専用回路が、連合野に形成されてゆく。これが、サル→人類と二段階に亘って、大脳連合野が飛躍的に発達していった理由である。
・構造認識は、大脳中枢の外識→内識をフル稼働させて照準を絞り込むことによって生み出される。その際、大脳中枢→大脳(専用回路)→連合野を何度も反復させて絞り込んでゆくが、そのサイクルを駆動させるのは連合野ではなく大脳中枢である。つまり、潜在思念が何らかの可能性を感取しない限り、連合野発でどれだけ追求しても、使える構造認識は生み出せない。
・また、外圧から遮断された学校での試験勉強等によって大半の潜在思念が封鎖されてしまった脳は、闘争照準力が極めて貧弱である。従って、照準がズレまくることになるが、狂った照準に基づいて観念回路だけで「何で?」を追求しても成果が0に終わる。文字脳・公式脳・暗記脳の人は、頭で考えるよりも、高速音読等で潜在思念を解除することの方が先決である。
(岡田淳三郎)

脳回路の仕組み10 人類の思考様式と構造認識
・人類が言葉を話しはじめて、60万年しか経っていないが、音声機能はすでにDNAに刻印されている。従って、それ以前に形成された観念機能(数十億本の神経回路)も、大きな大脳連合野(の神経回路)も、当然、DNAに刻印されている。
・それに対して、二元化や類型化等の把握機能は、DNA変異以前に、細胞質(おそらく中心体)に刻印されていると考えられるが、人類は、感覚機能の把握様式を踏襲しているだけのように見える。換言すれば、感覚機能の二元化や類型化を超えた、観念機能に固有の把握様式やその類型は未だ登場していないように見える。
・しかし、その萌芽はある。感覚機能の類型化を超えた観念機能に固有の類型認識として確立されてゆく可能性が高いと思われるのが、論理整合性に貫かれた構造思考と構造図解である。
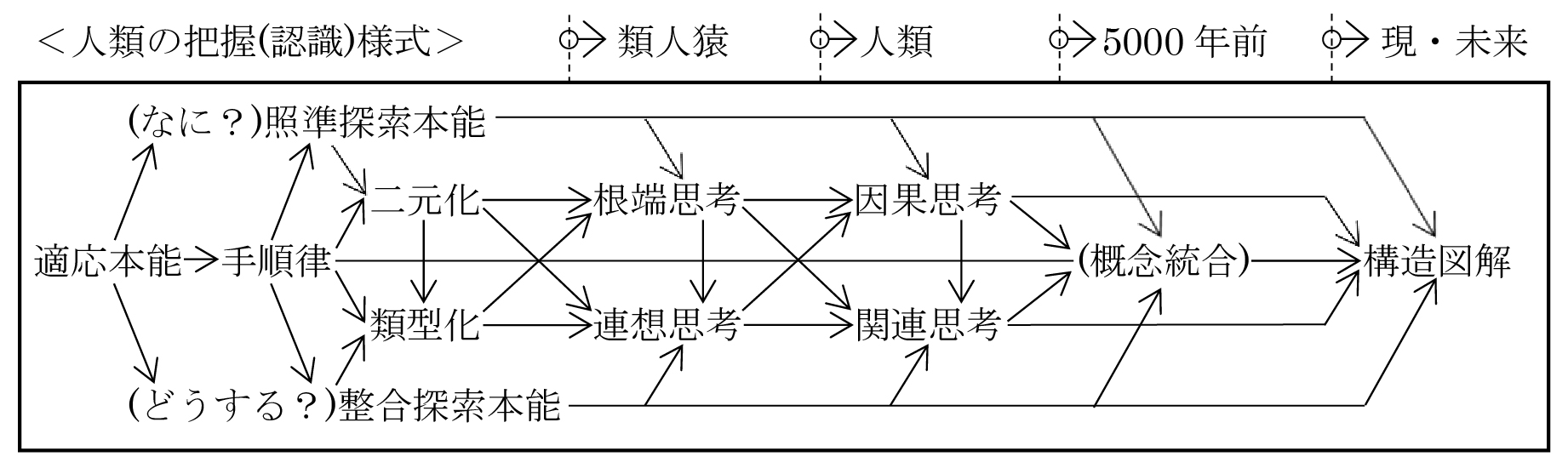
◎どうする⇒なんで?を生起させるのは、不整合感や危機感→探求核、および可能性展望→充足核の駆動物質。
◎人類は、未明対象の本質を把握する手法として、感覚機能の二元化と類型化を下敷きにして、根元と先端に二元化する根端思考と共通部がありそうな事象を探索する連想思考を発達させた。(共に感覚機能の応用)
◎どうする?や何?の探求は、必然的に根端や連想の各項の統合に向かうが、統合するためには各項の因果や関連を明らかにする必要がある。従って連合野は、因果思考や関連思考に先端収束していった。
◎しかし、部分的な因果や関連が分かってその部分が統合できても、全体との関連や位置は不明であり、その部分的認識が誤っている可能性もある。つまり、どうする?の答えになっていない=実現できない可能性がある。従って、因果思考や関連思考は、実現可能性を求めて、際限なく深化し、広域化してゆくことになる。それが実現思考=構造認識である。
(岡田淳三郎)
脳回路の仕組み9 生命の始源:中心体の仕組み
本稿は竹内美継の「中心小体論」を基に諸説を組み入れた仮説である。
1.ソマチッド(珪酸塩SiO4)と有機物と結合した蛋白粒が、螺旋状に伸びて微小管を形成する。この微小管が電磁波を受・発信するアンテナとなっている。
1’.細胞は、一次繊毛という数μmの小さな突起を持つが、この突起は微小管を繊毛膜で囲んだもの。この繊毛膜には、物理的・化学的信号を受信する受容体やイオンチャンネルが高密度に局在している。つまり、一次繊毛は、細胞が外部情報を感知するアンテナとして、細胞の維持や増殖などの第一義的な役割を担っている。
・微小管は弱伝導性で、電磁波を受信するシステムそのもの。八木アンテナは、伝導性の管がその長さに応じて特定の電波を受信するが、それとよく似た仕組みで、各波長の電磁波を受信している。この情報は、中心小体に集約していると思われる。(因みに、神経細胞の構造は、この一次繊毛の構造に類似している)
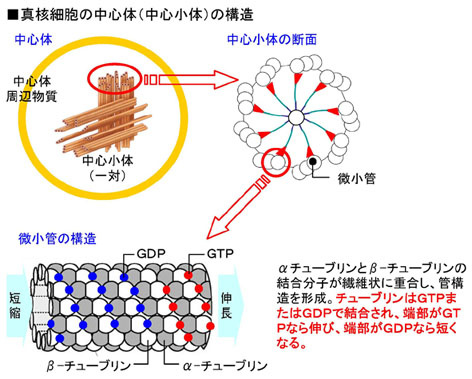
2.微小管はαチューブリンとβチューブリンが重合した二重螺旋の管構造。GTPやGDP等のヌクレオチドが糊のように働いて重合を可能にしている。(つまり、微小管にはRNA蛋白重合体が含まれている。)
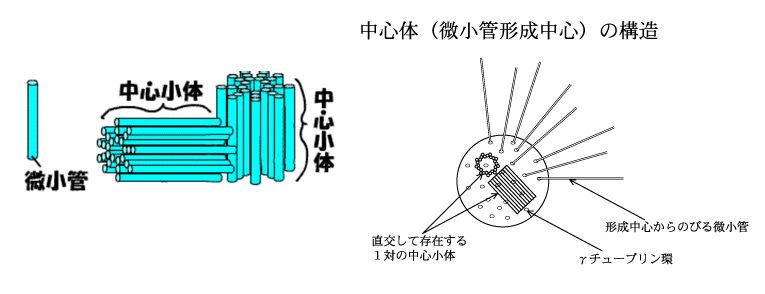
3.中心小体は、微小管が集合して形成された。
・微小管が集合して全体として一つの方向性を形成(中心体の祖先)
4.中心体は中心小体が2つ連なって構成されている。
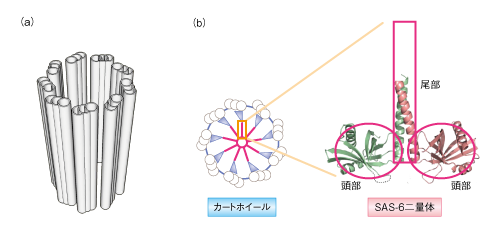
・L字型に直行する中心小体とそれを囲む顆粒状or繊維状の外周物質からなる。中心小体は直径200ミリミクロン、長さ700ミリミクロンの円筒。その構造は3連微小管が9本円筒状に並んだもので、その中心は一対の微小管。断面は、中心から伸びたタイヤのホイールの様な形状。
5.そのホイールを形成しているのは、SAS・6という蛋白質で、2つの球状の頭部と繊維状の尾部からなる二量体を形成している。この二量体どうしが頭部で結合することにより、9放射相称形という独特の形に会合する。
6.中心体が複製を作る機能を持ち、中心小体類と呼ばれる細菌(モネラ)が誕生。
・中心体は複製機能を持ち、始原生命体に最も近い生命体である。
・有糸分裂の際に紡錘体を形成し、細胞分裂の全体を統合している。 (中心小体が原核細胞と真核細胞を作った。DNAは中心小体の付帯装置に過ぎない。)
7.中心体周辺物質(γチューブリン)から中心体に向かって微小管が伸びているが、その微小管は、中心小体とは繋がっていない。
{kind=link}
(岡田淳三郎)
脳回路の仕組み8 把握様式の進化と脳容量の拡張
<把握様式の進化か、脳容量の拡張か?>
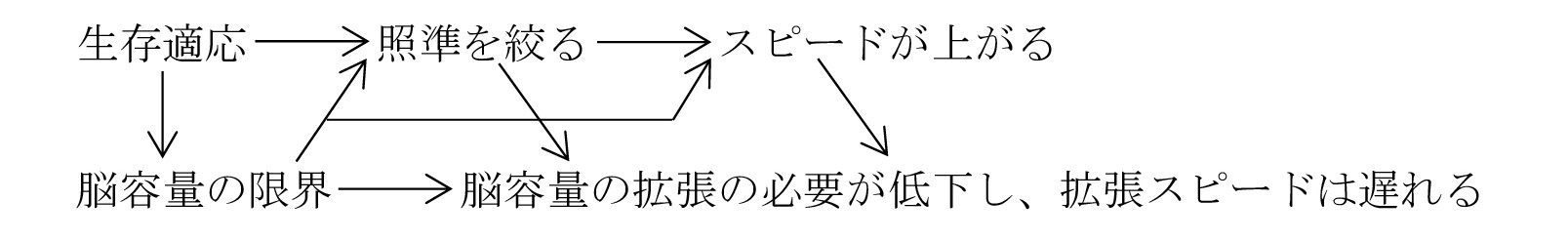
・脳容量の拡張にはDNA変異が必要で数万年はかかる。そこで、本能は最適手法たる照準化→類型化に収束。この照準化→類型化が先に細胞質(おそらく中心体)に刻印された。
・このことは、DNA進化に先行して、まず細胞質が進化することを明示している。考えてみれば、それは当然であって、卵子と精子の細胞質の全ては(精子の鞭毛とミトコンドリア以外)そっくり遺伝する。そして細胞質は必要発の駆動物質によって、DNA変異より遥かに容易に変異する。
・又、細胞質の変異は≒進化だが、DNAの変異はそのごく一部が進化に寄与するに過ぎない。更に、たとえDNA変異が中立(無方向)だとしても、その変異DNAの適・不敵を判断するのは駆動物質である。従って、進化の方向を規定しているのは、要・不要発の駆動物質であり、DNAではない(DNA進化は、その結果にすぎない)。
<視覚:形の類型化> 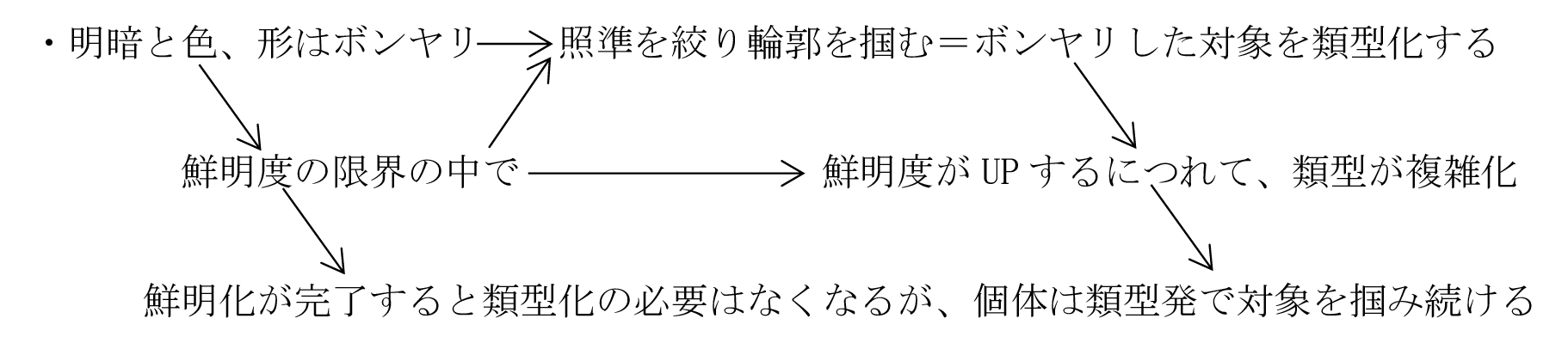
・鮮明度の上昇もDNA変異を待たねばならず、その前に形の類型化に向い、形の類型が先に細胞質(おそらく中心体)に刻印された。
・これは、鮮明度(解像度)を上げるのに必要な物質を増大させるDNA変異より、駆動物質発で把握機能を進化させる細胞質の変異の方が容易だからである。
・又、細胞質の変異は≒進化だが、DNAの変異は大半が不適応態で、ごく一部だけが進化に寄与するに過ぎない。更に、たとえDNA変異が中立(無方向)だとしても、その変異DNAの適・不適を判断するのは駆動物質である。従って、進化の方向を規定しているのは、要・不要発の駆動物質であり、DNAではない(DNA進化は、その結果にすぎない)。生物学者たちも、いい加減にDNA神話から脱却しないと、世間の笑い物になるだけだろう。
<赤ん坊:言葉の類型化>
・本能(細胞質)に刻印されている把握機能で言葉を類型化し、類型発で掴む→声に出す。
・音韻の類型認識→一音毎の類型認識→文法的な類型認識(語尾活用etc.)
・主要には1才前後によく聞く言葉を(本能に刻印された把握様式に基づいて)類型化している。ex.飲む、噛む、生む→死む。いない、しない→来(き)ない。
<人類の脳容量の急拡張>
・足の指が先祖返りして樹上で生きてゆけなくなった人類は、類型化etc.の把握機能を駆使してもとうてい対応できないような極限状態に陥った。
・そこで人類はひたすら「どうする?」をみんなで追求し続けるしかなかった。それが、チンパンジーの400ccから1400ccへと人類の脳容量が急拡張した理由である。言い換えると、人類の脳は「どうする?」を追求するためにある。その追求こそが、抽象化の極北に言葉(観念機能)をも生み出した。
・衝撃的な体験や映像は殆どが言葉化される。そして、一旦言葉化されると、その言葉を起点にして体験や映像を思い出すようになる。例えば、「幼児期の記憶」という言葉発でいくつかの体験や映像が思い起こされ、「3.11」という言葉でいくつかの出来事や映像が思い出される。
◎従って、言葉は様々な体験や映像の言葉化された(半)永久回路を飛躍的に増大させ、思い出し能力を飛躍的に上昇させる。それが、言葉(観念機能)を獲得して以降、人類の脳容量が飛躍的に拡張されると共に、知的能力が飛躍的に上昇した理由である。
・しかし、人類はその観念機能によって、約20万年前に極限状態を脱した。それ以降、人類の脳容量は増大していない。
そうなると、人類の進化は、他の動物と同様に、二元化→類型化→構造化という把握機能の進化に委ねられることになる。
・おそらく人類は、(現在既に登場しているが)論理不整合な文章(書き言葉)を脱して図解化に向かうだろう。
(岡田淳三郎)
脳回路の仕組み7 把握(認識)の二元化と類型化
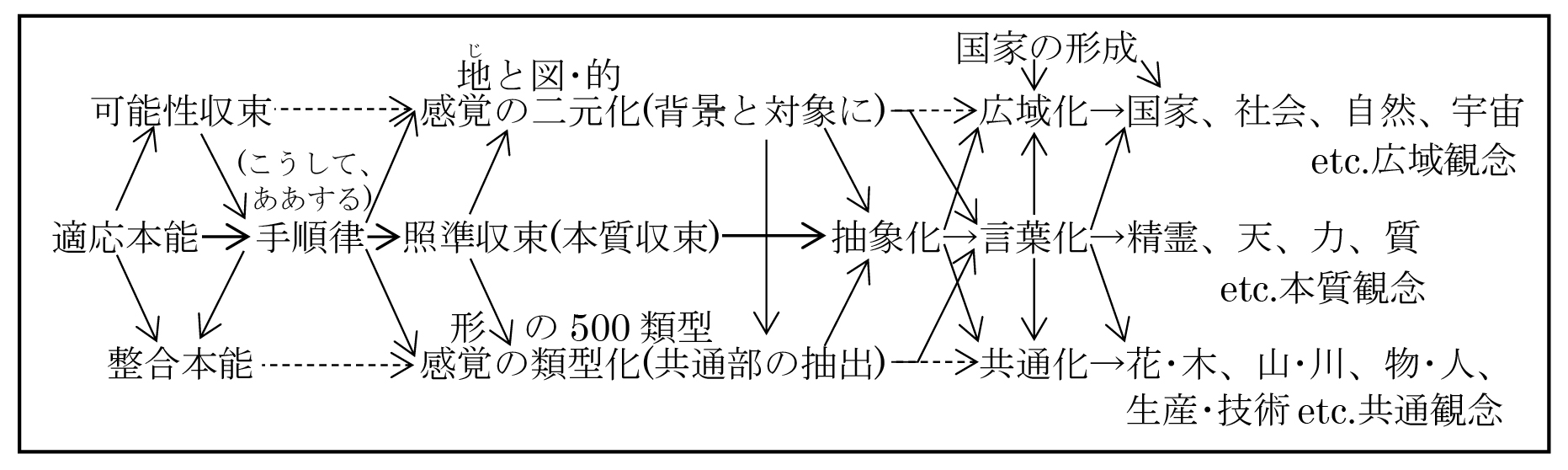
◎照準収束とは本質収束であり、それが手順律を下敷きにして感覚把握の二元化と類型化を生み出し、それらが抽象化を生み出した。この感覚機能の二元化と類型化および抽象化は、脊椎動物以来細胞質(おそらく中心体)に刻印されている。
◎意識は、類型に先端収束し、類型だけが深く意識に定着して先天機能となり、全ての対象を類型発で捉えるようになった。つまり、背景と対象に二元化したり、共通部を抽出して類型化したり、抽象化するのは、本能に備わった根元的な把握機能であり、カエルも、ネズミも、ネコも、無意識にそうしている。
◎本質収束とは最も重要な部分への収束であるが、その重要度を指示しているのは基盤神経や判断核etc.が放出する駆動物質である。
◎現実対象に対する照準収束→本質収束→抽象化の極北に生み出されたのが「精霊」を原観念とする観念機能である(100万~60万年前)。観念機能は仲間内で交信する必要から発声機能の発達を促し、それにつれて原観念は言葉として発信されていった。
◎原観念は共認機能の最先端に形成された窮極の抽象概念の像であり、それを音声化したものが、言葉=観念である。
(岡田淳三郎)
霊長類の大脳は、DNA(塩基配列)変化の伴わない進化形態である。
DNAの発見以降、生物の進化はDNAの変化に支配されていると言われていましたが、近年の分子生物学では、DNAに依らない進化(エピジェネティクス)≒ラマルクの進化論(要不要説 生物には「向上性」が内在していてこれが進化の要因と考えている。これが作用して、環境との相互作用の結果進化が起こる)
その代表例が人の大脳の進化であると思われる。
霊長類の大脳は、DNA(塩基配列)変化の伴わない進化形態である。
http://www.nibb.ac.jp/press/2013/12/19.htmlより
霊長類大脳皮質領野で特定の遺伝子のON/OFFが調節される仕組みの解明
大脳皮質は、ほ乳類の高次脳機能に中心的役割を担うものであり、霊長類、特にヒトで良く発達し、脳全体を覆うに至ります。大脳皮質はその場所によって異なる機能を持つことが知られており、それぞれ連合野、運動野、視覚野などの「領野」として区別されています。領野ごとの違いがどのように形成されるのかは、大変興味深いテーマですが、未だ多くの謎に包まれています。
基礎生物学研究所の脳生物学研究部門では、これまでに霊長類を用いて、脳の領野によって異なる発現パターンを示す遺伝子を発見してきました。それらの遺伝子は、例えば、連合野で発現し(ONになり)、視覚野では発現しない(OFFになる)、という発現の調節が行われていますが、領野の違いによって特定の遺伝子のONとOFFが調整される仕組みは、全く不明でした。
今回、畑克介研究員と山森哲雄教授らは、マカクザルの連合野ではONになり、視覚野ではOFFになる遺伝子の領野特異的な発現調節の仕組みの一端を明らかにしました。連合野特異的にONになる遺伝子のグループは、遺伝子発現を調節するプロモーター領域が高い割合でメチル化されていること、および、メチル化DNA結合タンパク質の一つとして知られるMBD4が連合野特異的に存在していることがわかりました。また、メチル化されたプロモーター領域にMBD4が結合することで、連合野特異的に遺伝子がONになることも明らかとなりました。これは霊長類の脳において、領野特異的な遺伝子のON/OFFの調節機構が明らかとなった初めての例です。
高度な認知機能を司る霊長類連合野に特異的に発現する遺伝子の発現機構はこれまで全く判っておらず、今回の成果は、今後の霊長類の大脳皮質の発達に関する研究と精神疾患の病因解明や治療等の研究につながる可能性が期待されます。この成果は、米国神経科学会誌Journal of Neuroscience(ジャーナルオブニューロサイエンス)2013年12月11日号にて発表され、「This Week in The Journal」として紹介されました。
[本研究の背景]
大脳皮質は「領野」と呼ばれる50程の区分に分けられ、それぞれの領野は、視覚や聴覚、運動機能などの機能を司ります。山森研究室では、霊長類の大脳皮質の領域特異的な機能分担や、脳の進化を探ることを目的として、マカクザル大脳皮質領野に特異的に発現する遺伝子の探索を行ってきました。霊長類大脳皮質の代表的領野である前頭連合野、側頭連合野、運動野、一次視覚野間で顕著な差のある遺伝子発現を比較検討し、RBP4遺伝子、PNMA5遺伝子、SLIT1遺伝子が連合野特異的に発現していること、HTR1B遺伝子、HTR2A遺伝子、FSTL1/OCC1遺伝子が一次視覚野特異的に発現していることを明らかにしてきました。
霊長類以外(例えばネズミやウサギやフェレットなど)では、これらの遺伝子は、領野特異的な発現を示しません。このことは、霊長類大脳皮質の領野特異的遺伝子の発現メカニズムは霊長類への進化の過程で備わったものであり、この機構を明らかにすることは、霊長類の脳の発達や進化のメカニズムの解明に重要な知見をもたらすものと期待されます。これら領野特異的遺伝子の発現パターンが各グループで良く似ていることから、共通の発現制御機構があると推測してその解明を目指しました。
[本研究の内容]
研究グループは、連合野において特異的に発現する遺伝子のグループ(RBP4遺伝子, PNMA5遺伝子, SLIT1遺伝子)と、一次視覚野において特異的に発現する遺伝子のグループ(HTR1B遺伝子, HTR2A遺伝子, FSTL1遺伝子)の、プロモーター領域のDNAメチル化の程度を解析しました。その結果、連合野に特異的な発現をする遺伝子グループのプロモーター領域は高度にメチル化されているが、一次視覚野に特異的な発現をする遺伝子グループのプロモーター領域はメチル化の程度が低いことを発見しました。
さらに、メチル化DNA結合タンパク質の一つとして知られるMBD4の遺伝子発現が連合野特異的遺伝子と非常に似た発現を示すことがわかりました。
また、メチル化DNA結合タンパク質MBD4が連合野特異的遺伝子の高度にメチル化されたプロモーターに結合し、その結果として連合野に特異的な遺伝子発現を促すことが明らかとなりました。DNAのメチル化による遺伝子発現の制御は、DNAの塩基配列の変化を伴わないエピジェネティクスな機構として、生物の多様な表現型の形成や、種々の発達障害、がん化などに関わることが近年明らかになってきています。本研究は、霊長類の脳において、領野特異的な遺伝子のON/OFFの調節機構が明らかとなった初めての例であり、その調節がDNAのメチル化によるエピジェネティクスな機構でコントロールされていることを示しました。
[今後の展開]
高度な認知機能を司る霊長類連合野に特異的に発現する遺伝子の発現機構はこれまで全く判っておらず、今回の成果は、今後、霊長類の大脳皮質の発達に関する研究、精神疾患の病因解明や治療等につながる可能性があります。
以上です。
脳回路の仕組み6 成績圧力と文字脳・公式脳
・誰もが文字を読めるようになって、たった150年で言語能力がドン底にまで劣化してしまった原因は、「そもそも文字は潜在思念とのつながりが貧弱である」という文字の根本欠陥だけにあるのではない。
・私権社会に特有の私権の強制圧力→学校の強制圧力→勉強の強制圧力が、子供の意欲や追求心を封鎖してしまうという現実の圧力の方が、はるかに大きな原因として在る。
・貧困が消滅して、すでに50年。貧困の圧力に基づく私権の強制圧力(私権を獲得しなければ生きていけないという否も応もない強制圧力)はとことん衰弱し、代わって仲間充足と追求充足が新しい活力源として、すでに顕現している。従って、今や私権の強制圧力を土台とする「いい生活⇒いい大学⇒いい成績」では全く意欲が湧かないどころか、その強制圧力が仲間充足・追求充足という活力源を封じ込めるので、子どもたちの肉体的な拒絶反応を引き起こし、それが年を追って深刻な状態になってきている。
・しかし、嫌だから引きこもるでは答えにならない。それでは敗け犬にしかならない。実は目先の成績に囚われている親→子ほど、より強く時代遅れの私権の強制圧力に囚われており、それ故、子供の肉体的な拒絶反応はより深刻な状態にあるので、何かのきっかけで簡単に引きこもり状態に陥る。
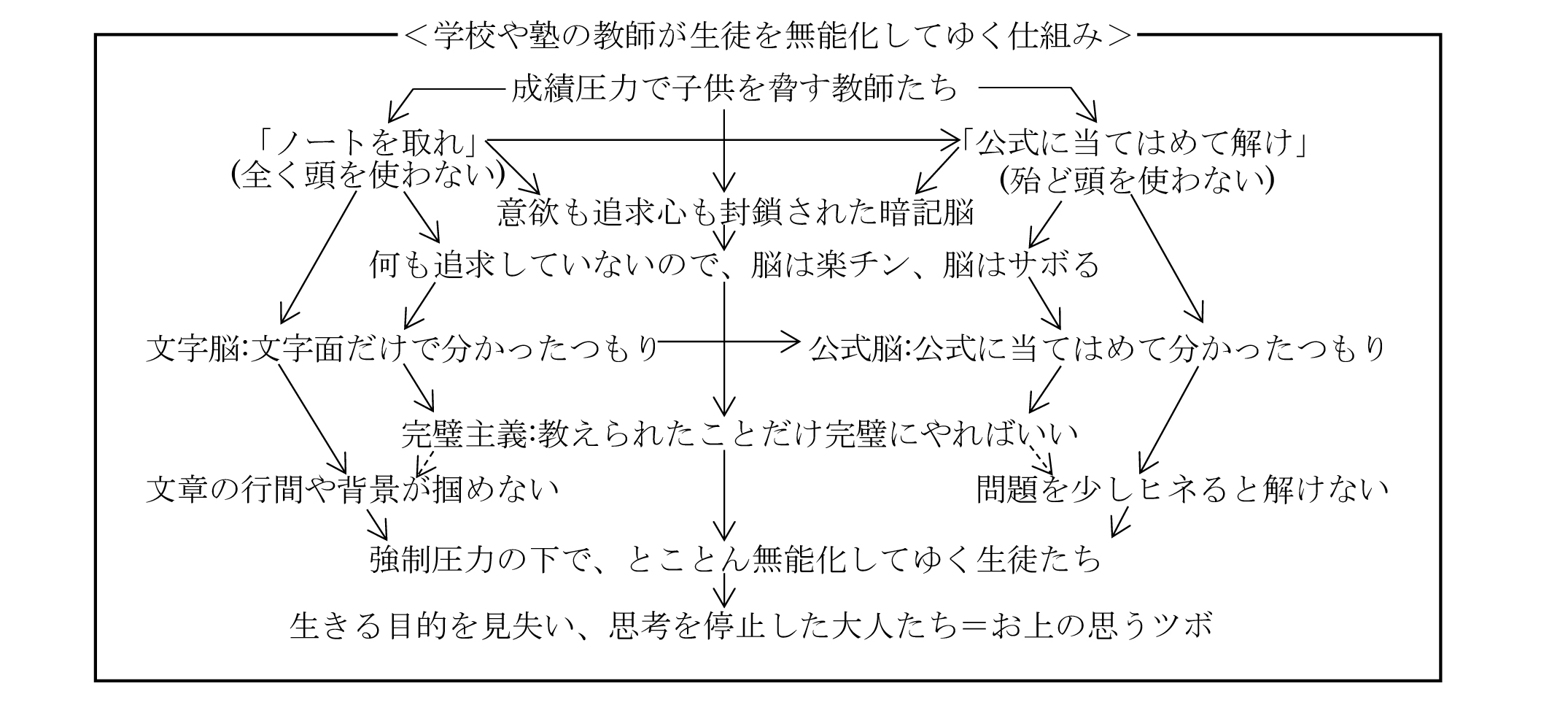
・私権社会の共認形成は、窮乏(きゅうぼう)圧力であれ、利益誘導であれ、あるいはいじめ集団の親分・子分であれ、学校の教師の脅しであれ、国家の権力であれ、全て私権の強制圧力を基盤とする脅し→騙しによって成り立っている。脅されると、脳の危機逃避の回路が強く作動し、その逃避先として与えられた所に簡単に収束するからである。
・学校について言えば、私権の強制圧力を背景とする成績圧力で親と子供を脅して、「ノートを取れ」「公式で解け」と誘導し、そうして意欲も追求心も奪い取り、ひたすら暗記脳・文字脳・公式脳に染め上げてゆく。その結果が、
意欲も追求心も失い、思考停止に陥った子供たち→大人たちである。
・しかし、今や暗記脳の試験エリートなど、社会に出れば使い物にならない。だから、これは大いなる騙しである。しかし、それが騙しであることに気付く人は少ない。親も子供も、大半が学校の脅しに騙されている。だから、「脅しに騙されるな」、「今や学校は、国民を無能化する装置でしかない」ことを、声を大にして周りに訴える必要がある。
・しかし、現に学校の強制圧力に晒されている生徒たちは、どうしたら良い?
まずは、脅しに騙されないこと=目先の成績から脱却すること、そして、本来の追求心を解放して強制圧力を突き抜けてしまうこと。
・追求力さえ身につけば、(1年ぐらいで)追求力が成績圧力を突き抜けて、実は簡単に成績も上がってゆく。もっとも、学校の成績を上げる気が全くない生徒は、別に上げなくても追求力さえ身に付ければそれで良いが。

・自分の頭で追求して得た認識は、その答えに達するまでに関連する認識群を何度も反復しているので、3~5回ほど反復するだけで永久回路として定着する。
・教科書etc.で与えられた認識は、内識と殆ど繋がっていないので、100回以上反復しないと定着しない。しかも、100回反復しても半永久回路にしかならず、1年後、3年後、10年後には曖昧にしか思い出せない。
・言語能力を形成する秘訣は、赤ん坊が言語を習得する過程に凝縮されている。即ち、聞いて、真似して、しゃべるを繰り返すこと。それが最も効率的な言葉の習得法である。国語の成績を上げる秘訣も同様で、ひたすら相手の表情や口元を真似して(文字は見ずに)高速で音読すること。そうすれば封鎖された潜在思念が解放されて能力が上昇する。
・又、英語などの新しい言葉を吸収する場合も、赤ん坊と同様にゼロから直結回路を形成してゆくのが最も効率的である。この場合、既存の日本語を媒介にしていると、スピードが遅すぎて会話についてゆけないだけではなく、直結回路の形成が疎かになり、捨象されてしまう。従って、あくまでゼロから(聞いて、真似して、しゃべるを繰り返して)英語脳を形成してゆくことが、外国語を吸収する最短の道である。
(岡田淳三郎)
長期記憶形成に必須な分子メカニズムを特定
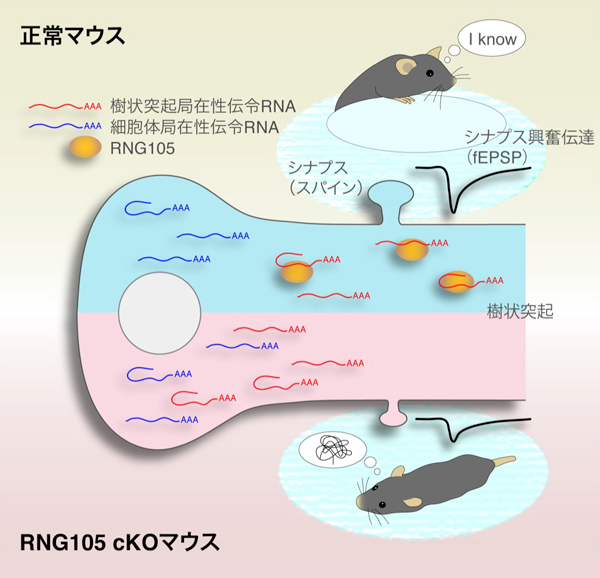
長期にわたる記憶形成するためには、ニューロンの樹状突起を肥大化しシナプスと呼ばれるつなぎ目と繋がりやすくする必要があります。そのメカニズムの一端を明らかにした研究報告が有りましたので転載します。
基礎生物学研究所 http://www.nibb.ac.jp/press/2017/11/21.html 2017年11月
長期記憶形成に必須な分子メカニズムを特定 ~タンパク質の設計図を神経樹状突起へ局在化させる因子が不可欠~
(前略)
我々が物事を覚える際に、数時間、数日間、あるいは数年間という長期に渡る記憶を形成するためには、脳内でのタンパク質合成が不可欠であることが知られています。しかし、そのタンパク質合成の分子メカニズムは四半世紀に渡って未解明の問題として残されていました。神経細胞におけるタンパク質合成には、細胞全体で起こる一般的な制御と、神経細胞間の興奮伝達を仲介する樹状突起上のシナプスの近傍でのみ起こる局所的な制御とが存在します。本研究では、後者の局所的制御に関わる因子「RNG105」に着目し、マウスを用いて、RNG105の欠損が長期記憶や伝令RNAに及ぼす影響について解析を行いました。その結果、RNG105欠損マウスでは、特定の伝令RNAの樹状突起への局在化が低下し、長期記憶が著しく低下することが明らかになりました。これら特定の伝令RNAをもとに合成されるタンパク質には、シナプスでの興奮伝達を担う「AMPA受容体」を制御するタンパク質が多く含まれていました。RNG105欠損マウスでは、このAMPA受容体が上手く機能できず、シナプスを介した興奮伝達も著しく低下しました。以上の結果から、RNG105によって伝令RNAを樹状突起に局在化させ、その伝令RNAをもとにしてAMPA受容体制御タンパク質の合成を樹状突起上のシナプス近傍で起こすことが、シナプスの正常な興奮伝達、ひいては長期記憶の形成に不可欠であるという新たな分子メカニズムを提唱しました。
本研究成果は、2017年11月21日付けで英国オンライン科学誌eLifeに掲載されます。
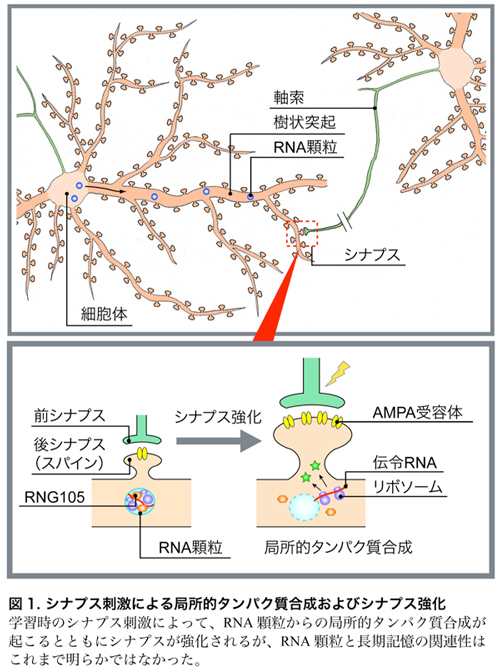
【背景】
私たちの脳を構成する神経細胞(ニューロン)は、2種類の長い突起(軸索・樹状突起)を持ち、あるニューロンの軸索と異なるニューロンの樹状突起がシナプスと呼ばれるつなぎ目で結合しています(図1)。長期記憶の形成には、樹状突起上にある後シナプス(スパイン)を肥大させ、そこにAMPA受容体*1を増やすという「シナプス強化」が不可欠です(図1)。このシナプス強化のためには、樹状突起のスパイン近傍で「局所的タンパク質合成」が起こることが重要だと考えられています(図1)。局所的タンパク質合成が起こるためには、タンパク質合成の設計図である伝令RNAが、「RNA顆粒」と呼ばれる複合体により細胞体から樹状突起へと輸送されることが必要です(図1)。しかし、RNA顆粒の働きが実際に長期記憶に必要かどうかは、長年の間、未解明の問題として残されていました。
本研究では、RNA顆粒の主要な構成因子“RNG105”(別名Caprin1)に着目しました。これまで、RNG105を恒常的に欠損したマウスは生後まもなく死亡するため、成体マウスにおけるRNG105の機能は不明のままでした。そこで本研究では、RNG105を胎仔期後期以降、脳で欠損させたRNG105コンディショナル欠損 (cKO) マウスを作製し、成体マウスを得ることに成功しました。このマウスを用いて、RNG105が長期記憶に与える影響、およびその基盤となる分子メカニズムの解明を目指して研究を行いました。
【研究成果】
・RNG105コンディショナル欠損 (cKO) マウスでは長期記憶が低下する
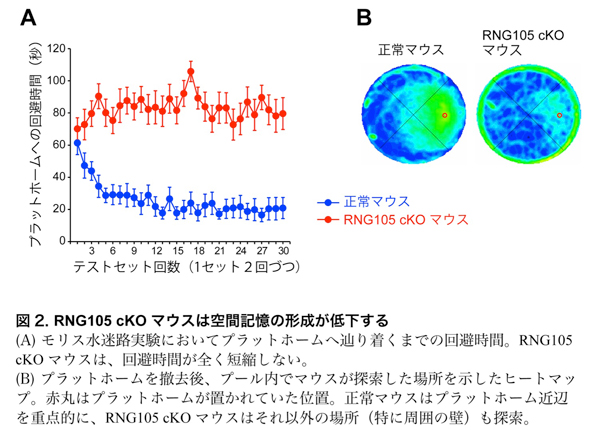
RNG105 cKOマウスの学習・記憶能力について、モリス水迷路テストおよび条件付け文脈学習テストにより測定を行いました。モリス水迷路テストは、円形プールの特定の場所にマウスが回避できるプラットホームを隠して置き、その回避場所を数日かけて覚える長期記憶テストです。正常マウスはテストを重ねるごとにプラットホームの位置を学習して記憶したのに対し、RNG105 cKOマウスはテスト回数を重ねてもほぼ全く記憶ができませんでした(図2)。
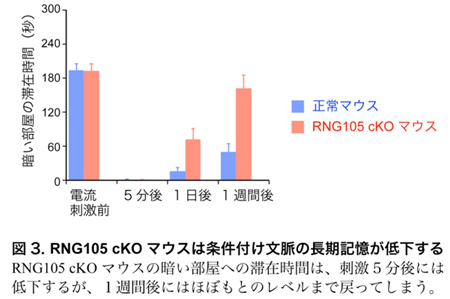
条件付け文脈学習テストでは、明るい部屋と暗い部屋とを自由に行き来できる装置を用います。通常、マウスは暗い場所を好み、そこに長く滞在します。しかし、暗い部屋に入った際に弱い電流を経験すると、マウスは嫌悪の経験を記憶し、その後は電流が流れなくても暗い部屋の滞在時間が減少します。この暗い部屋の滞在時間を計測した結果、RNG105 cKOマウスは5分間の短期記憶は正常に形成されるのに対し、1日から1週間に渡る長期記憶の形成は著しく障害されていることが明らかになりました(図3)。
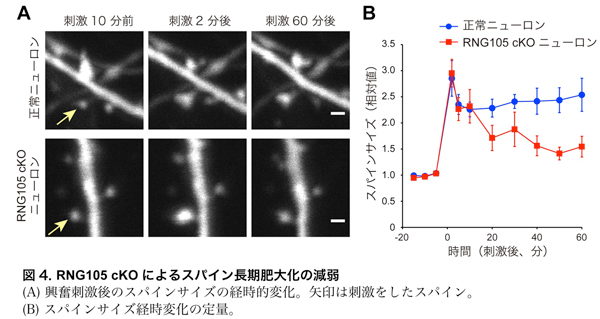
・RNG105 cKOニューロンではシナプス強化 (スパイン肥大) が減弱する
RNG105 cKOのシナプス強化に対する影響を調べるために、興奮刺激を受けた際のスパインの肥大化(図1)を計測しました。正常ニューロンでは刺激によりスパインが肥大し、そのサイズは刺激後1時間経っても維持されました。一方、RNG105 cKOニューロンでは刺激によりスパインは一度肥大したものの、時間経過とともに次第に縮小しました(図4)。よって、RNG105は長期的なスパイン肥大化(シナプス強化)に必要であることが示されました。
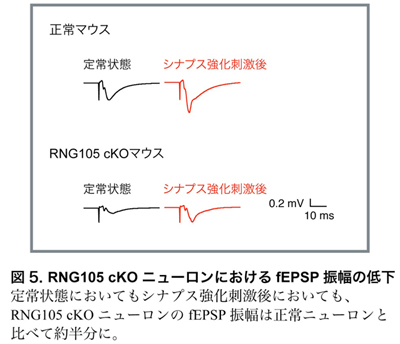
・RNG105 cKOニューロンではシナプス興奮伝達が低下する
次に、シナプス興奮伝達の指標となる興奮性シナプス後電位(fEPSP)の測定を行いました。その結果、RNG105 cKOマウスは正常マウスと比較し、fEPSPの大きさが約半分に低下しました(図5)。このことは、RNG105 cKOマウスでは、シナプスの興奮伝達を担うAMPA受容体が上手く機能できず、興奮伝達が低下していることを示しました。
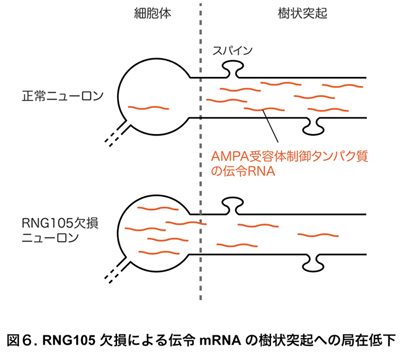
・RNG105 cKOニューロンでは樹状突起への伝令RNAの局在化が低下する
以上のようなRNG105 cKOマウスの表現型の基盤となる分子メカニズムを解明するため、RNG105 cKOが伝令RNAに及ぼす影響を解析しました。その結果、通常ではニューロンの樹状突起へ局在化する特定の伝令RNA群が、RNG105 cKOニューロンの樹状突起では局在化が低下していることを見出しました(図6)。そのような伝令RNA群の中には、AMPA受容体を制御するタンパク質の設計図である伝令RNAが多数含まれていました。以上のことから、RNG105 cKOニューロンでは、樹状突起上のスパイン付近へ本来供給されるべき伝令RNAが不足し、それをもとに合成されるAMPA受容体制御タンパク質もスパイン付近では不足していると考えられました。
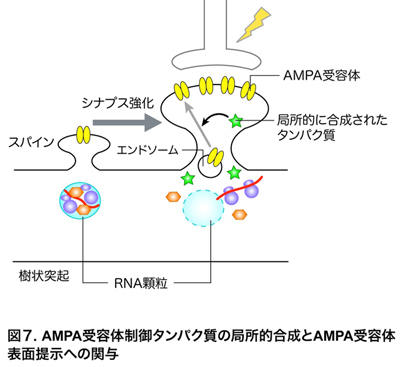
・RNG105の欠損は樹状突起におけるAMPA受容体の表面提示制御を低下させる
AMPA受容体制御タンパク質は、樹状突起内のエンドソームに隔離されたAMPA受容体を細胞表面に提示し、そこにつなぎ止めることによって、シナプス強化に関わることが知られています(図7)。そこで、AMPA受容体が樹状突起の表面にどのくらい提示されているかを定量解析しました。その結果、RNG105欠損ニューロンでは、樹状突起の表面にAMPA受容体を提示する制御機構が上手く機能せず、正常ニューロンに比べて表面のAMPA受容体が減少していることが明らかになりました。
以上の知見をまとめると、RNG105によって特定の伝令RNAが樹状突起に局在化し、樹状突起上のシナプス近傍でその伝令RNAをもとにしてAMPA受容体制御タンパク質を合成することが、シナプス強化、すなわち、スパインを肥大させてその表面に多くのAMPA受容体を提示させるのに必要だと考えられます。本研究は、このメカニズムにともなう分子が、シナプス興奮伝達ひいては長期記憶の形成のために必要であるという新たなモデルを提唱しました。
【本研究の意義と今後の展開】
長期記憶形成にはタンパク質合成が必要であることが知られていたものの、両者をつなぐ分子メカニズムはこれまで明確に示されていませんでした。本研究は、RNA顆粒の構成因子であるRNG105が長期記憶の形成に不可欠であることを明らかにしました。さらに、その基盤となる分子メカニズムとして、RNG105は樹状突起への特定の伝令RNAの局在化を制御し、AMPA受容体の樹状突起における表面提示制御に関与することを示しました。
記憶は生物の生存や精神活動の基盤となる脳機能です。記憶の障害は様々な精神神経疾患とも関連しています。また、RNA顆粒の構成因子には、筋委縮性側索硬化症(ALS)等の神経変性疾患の他、精神遅滞症、自閉症スペクトラム(ASD)などの原因因子が数多く含まれています。RNG105の変異も、ASD様の行動を引き起こす原因の一つです。したがって、RNG105およびRNG105が樹状突起へ輸送する伝令RNA群は臨床的なマーカーや創薬のターゲットとなりうる可能性を有しており、本研究の成果は、記憶形成の分子メカニズムのさらなる解明に加え、臨床・創薬分野において役立つことが期待されます。
【用語説明】
*1AMPA受容体:興奮性神経伝達物質であるグルタミン酸の受容体の一種。中枢神経系のシナプス興奮伝達は主にこの受容体が担い、学習・記憶に必須の役割を果たしている。
脳内にある、やる気のスイッチを発見
脳内でやる気を起こす部位を見つけたとする論文が有りましたので転載します。
、2017年2月1日に総合科学雑誌であるNature Communicationsに掲載されたものです。
http://www.nips.ac.jp/release/2017/02/post_335.html
_____________________________________________
|
脳内にある、やる気のスイッチを発見
|
1.研究の背景と概要
認知症などの神経変性疾患、脳血管障害や脳外傷などの脳の障害では、いずれも高い頻度で意欲障害が認められます。いわゆる「やる気がない」という症状であり、リハビリテーションの阻害因子として患者さん本人のQOL(quality of life)を低下させるのみならず、介護者の意欲を削ぐ要因にもなります。うつ病の意欲障害には、抗うつ薬という治療の選択肢がありますが、損傷脳の意欲障害にはどの薬が有効で、何が無効かなど治療薬選択について全く分かっていません。その一つの要因として、損傷脳の意欲障害がどのようなメカニズムによって発生するのか全く分かっていないので、候補薬さえも挙げられない状況です。
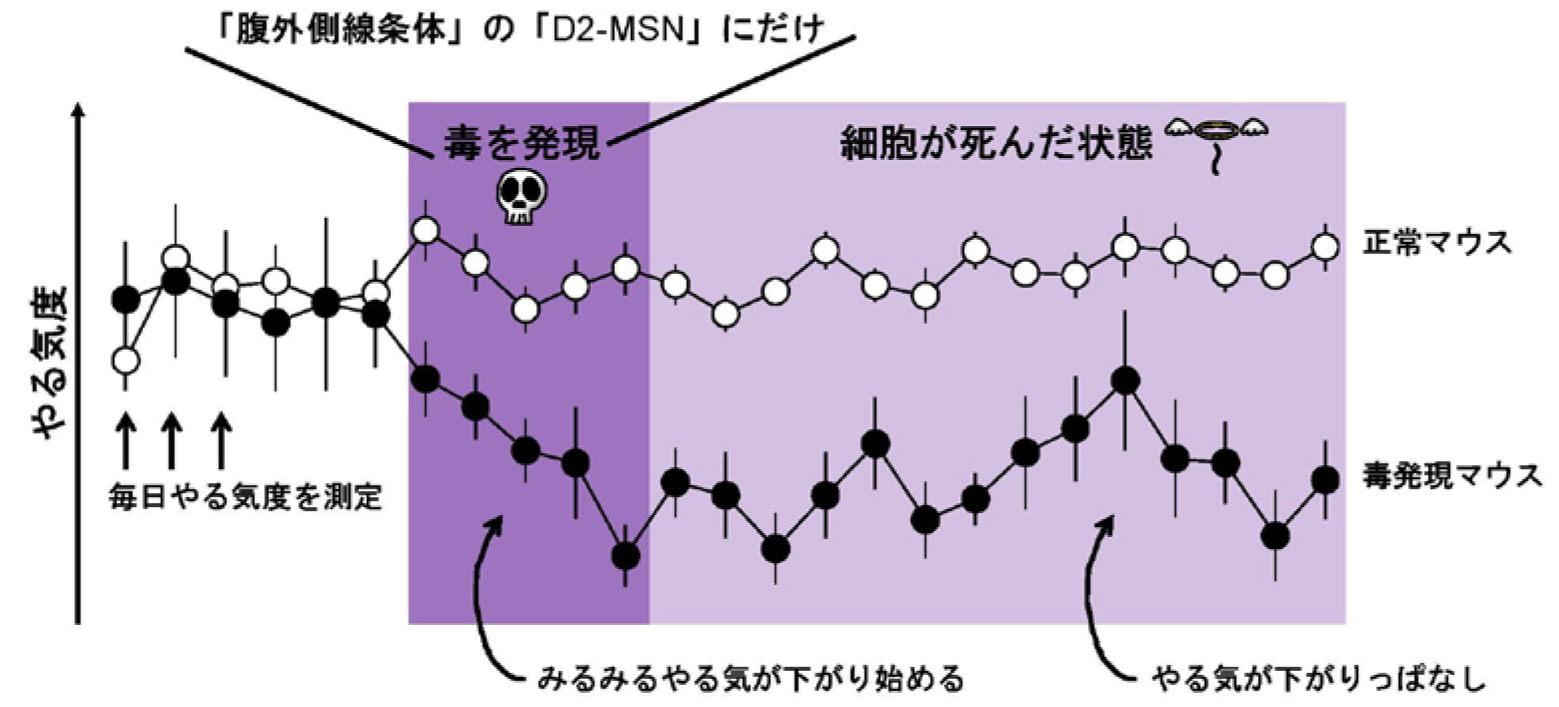
研究グループは、脳の特定部位である線条体(注1)の損傷によって意欲障害を起こす頻度が高い臨床結果を参考にして、線条体を構成する一つの細胞集団、ドパミン受容体2型陽性中型有棘ニューロン(注2)(以下D2-MSN)に注目しました。実験者が任意のタイミングでD2-MSNを除去することができる遺伝子改変マウスを作出し、意欲評価の実験を行いました。マウスの意欲の評価には比率累進課題(注3)と呼ばれる餌報酬を用いた行動実験を用いました。
あらかじめマウスに課題を学習させておき、マウスの意欲レベルを調べます。その後、D2-MSNだけに神経毒を発現させて徐々に細胞死させます。もしもD2-MSNが意欲行動をコントロールするならば、D2-MSNの細胞死によって、マウスの意欲レベルは下がるはずです。また、意欲の低下が線条体のどの部位の損傷で、どの程度の損傷の大きさで起こるのかわかるはずです。
研究の結果、線条体の腹外側(注4)の障害で、かつ、その領域のわずか17%の細胞死によって意欲障害が起こることが分かりました(図1)。研究グループは、神経毒以外の方法、すなわちオプトジェネティクス(注5)によるD2-MSNの機能抑制、オプトジェネティクスによるD2-MSNの破壊という2つの異なる方法によっても、腹外側線条体のD2-MSNが意欲行動に必須であることを見出しました。
図1
2.研究の成果と意義・今後の展開
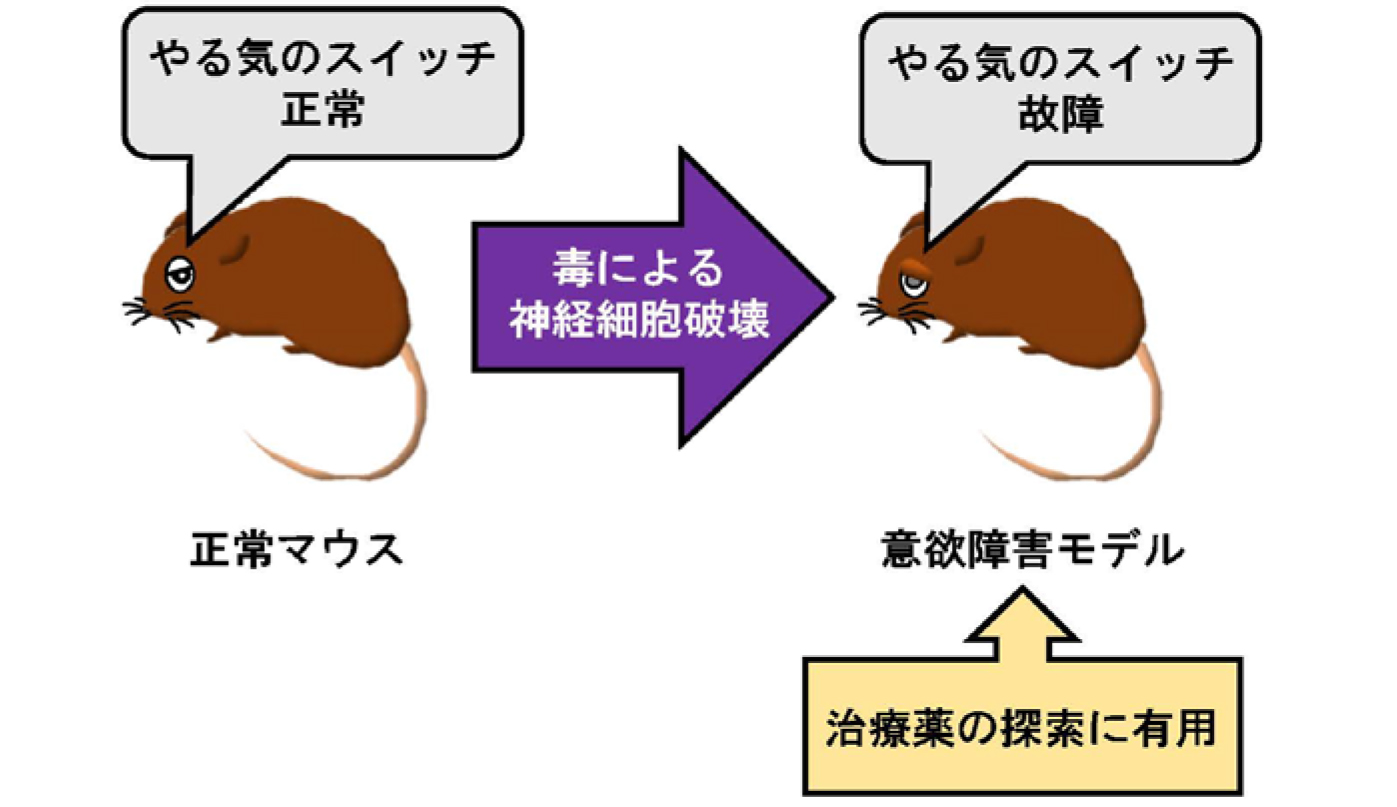
動物を使った意欲の研究では、おいしい餌を報酬とする場合と、覚せい剤のような依存性薬物を報酬とする場合があります。依存性薬物を希求する意欲の責任脳部位として線条体の腹内側部が知られていましたが、おいしい餌のような生理的な欲求に対する意欲の責任脳部位は分かっていませんでした。本研究によって、その責任脳部位が線条体腹外側部であること、中でもD2-MSNが意欲の制御に働いていることが明らかになりました。他にもいくつかの部位が「やる気」を生むのに必要であると想像されていますが、本研究によって初めて、やる気を維持する脳部位・細胞種を明確に示しました。
損傷脳の意欲障害のモデル動物が樹立できましたので、今後はこのモデル動物を用いて、意欲障害を改善する薬剤を探索することができます(図2)。
図2
【用語解説】
(注1)大脳基底核・線条体:運動制御や報酬を計算する脳部位で、大脳皮質に囲まれた脳深部(基底)にある。いくつかの脳部位から成る。線条体はそのうちの一つ。
(注2)ドパミン受容体2型陽性中型有棘ニューロン:線条体から情報を外に送る神経に2種類あり、一つがドパミン受容体1型陽性中型有棘ニューロンで、もう一つがドパミン受容体2型陽性中型有棘ニューロンである。ドパミン受容体2型陽性中型有棘ニューロンはこれまで、意欲を押さえる役割を果たすと考えられてきた。
(注3)比率累進課題:1個の餌報酬を得るのに、課されるレバー押し回数が増えていく課題。はじめは1回レバーを押せば1個の餌をもらえるが、2個目の餌を得るのには2回レバーを、3個目の餌を得るには4回、4個目の餌を得るには6回という具合にレバーを押す回数が増えていく。たとえば14個目の餌を1つ得るのに95回レバーを押す必要がある。
課題中のマウスはいずれ餌を得ることをあきらめる。諦めるまで頑張った回数が意欲の程度を表す。意欲が下がると諦めるまでのレバー押し回数が減る。
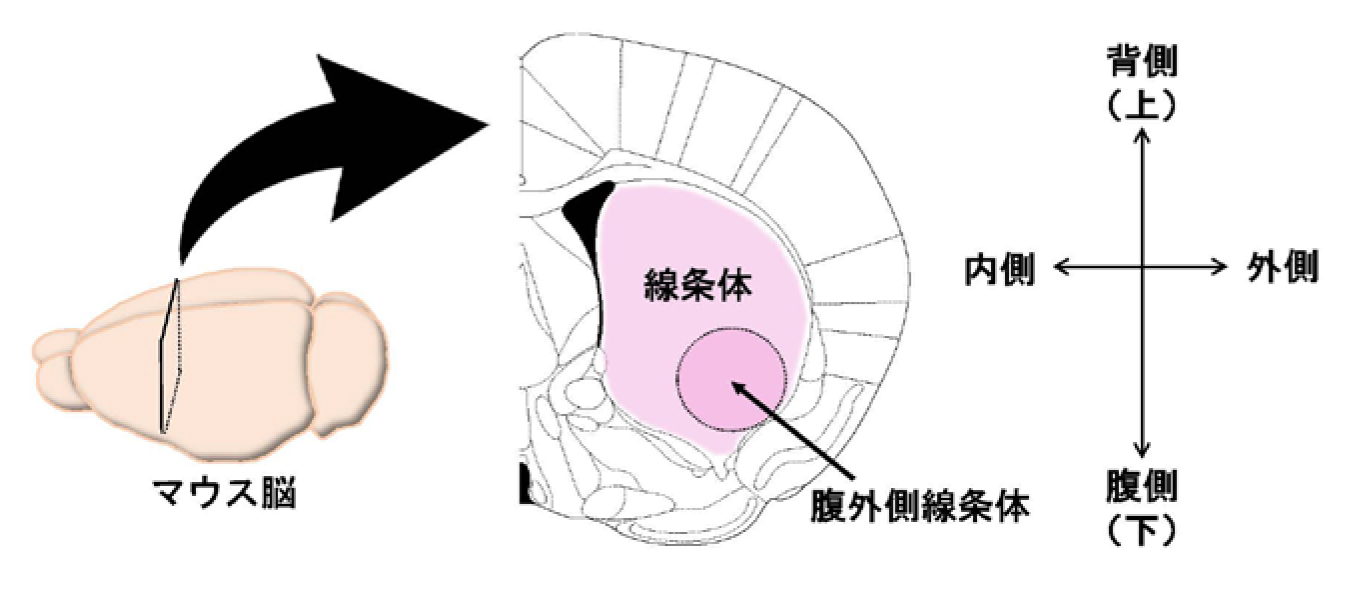
(注4)線条体腹外側:線条体の下部(腹側)かつ外側の領域のこと。一方で、線条体腹内側は、線条体の下部で内側の領域(図3)。
図3
(注5)オプトジェネティクス:光を用いて特定の細胞集団の活動を操作する技術。このケースでは、ドパミン受容体2型陽性中型有棘ニューロンの活動を光で操作した。
ダーウィンの進化論が崩壊:進化は不連続に起こる。ミッシングリングは存在しない。
学校の教科書で、生物の進化は「ダーウィンの進化論(連続的に徐々に進化)」と記載されている。ただ人類の進化には、ミッシングリングという謎(猿から類人猿そして現生人類をつなぐ中間体の化石が見つからない)が有ると教えられてきた。
しかし近年の遺伝子検査により生物の進化は「急激に進み新種が誕生し、いったん誕生した新種は滅びるまでほとんど変化しない」ようである。
そもそも中間体というのは存在せず、昆虫は太古から変化してない。又シーラカンスもワニもライオンも何十万年も形態は変わってない。進化は不連続に突然起こるのではないかと考えられる。
生物の主要な構成物質である水も周囲の環境(エネルギーの授受で)で個体・液体・蒸気・プラズマ(水素と酸素)状態へ瞬時に変化する。そして素粒子さえも不連続に誕生したり消滅したりしている。
とすると、生物も、外部環境に適応できない種は滅亡し、先祖返りして適応した種を作り出す戦略(≒進化は不連続に起こる)を採っていると思える。
[特報]ダーウィンの進化論が崩壊:かつてない大規模な生物種の遺伝子検査により「ヒトを含む地球の生物種の90%以上は、地上に現れたのがこの20万年以内」だと結論される。つまり、ほぼすべての生物は「進化してきていない」
との研究発表がありましたので転載します。
IN DEEP https://ameblo.jp/glamzanzedeus/entry-12420093747.html
________________________________
(前略)
これはつまり、この地球の生物の 90%以上は「それ以前への遺伝子的なつながりがない」ということでもあり、もっといえば、
・地球のほとんどの生物は 20万年前以降に「この世に現れた」
これがどういう意味かといいますと・・・。たとえば・・・「現行の科学で言われている人類誕生までの地球の歴史」というものは下のようにされています。
46億年前から始まり、35億年前くらいの最初の生物が誕生し、そこから「徐々に」進化してきた……というものです。
しかし、今回の大調査の結果からわかることは、
「徐々に」進化していないということなのです。
つまり、20万年より前の部分は、「現在の地球の生物とほとんど関係ない」としか言いようがないのです。
とにかく、ほぼすべての生物種が 10万年から 20万年前に地球に登場しているという可能性が極めて強くなったのです。
生物の全面的な大規模遺伝子調査により、生物進化の新しい側面が明らかに
これは、アメリカ政府が運営する遺伝子データバンク(GenBank)にある、世界中から数百人の科学者たちによって集められた 10万種の生物種の DNA と、500万の遺伝子断片である DNA バーコードと呼ばれるマーカーが徹底的に調べ尽くされたのだ。
それを行い、その「結果」を報告したのは、米ニューヨーク・ロックフェラー大学のマーク・ストークル(Mark Stoeckle)氏と、スイス・バーゼル大学のデビッド・タラー(David Thaler)氏であり、共同でその内容が発表された。
そして、その内容は「生物の進化がどのように展開されたか」についてのこれまでの定説を揺らがせるものだったのだ。
定説とは何か?
現在の生物学の教科書では、たとえば、アリでもネズミでもヒトでもいいのだが、大規模な個体群を持つ生物種は時間が経過するほど遺伝的多様性が増すとされている。このように時間の経過と共に、生物が進化してきたというのが定説だ。
しかし、それは本当なのだろうか?
その問いに対して、今回の研究の主任著者であるマーク・ストークル氏は次のように述べた。
「いいえ、それは違います」
ストークル氏は、地球上に住む 76億人のヒトも、5億羽生息しているスズメも、あるいは、10万羽生息しているシギたちも、その遺伝的多様性は「ほぼ同じくらいなのです」と AFP に語った。
動物には 2種類の DNA がある。核 DNA とミトコンドリア DNA だ。しかし、すべての動物はミトコンドリア内に DNA を持っている。
ミトコンドリアは、細胞からのエネルギーを食物から細胞が使用できる形に変換する各細胞内の小さな構造体だ。生物の種と種の間で大きく異なる可能性のある核 DNA 遺伝子とは異なり、ミトコンドリア DNAにはすべての動物が持つ共通の DNA 配列が存在する。
この共通の DNA 配列が比較のための基盤を提供するのだ。
カナダの分子生物学者であるポール・エイバート(Paul Hebert)氏は、2002年頃に「 DNA バーコード」という用語を作り出し、COI 遺伝子を解析することで種を同定する方法を描いた。
そして、研究者が目にしたものは、いわゆる「中立」な遺伝子変異にばらつきがないことだ。この中立突然変異が、互いにどれほど類似してるかは樹木の年輪を見るようなもので、これにより一つの種のおおよその年齢が明らかになる。知られているところでは、最後の地球での大量絶滅事象は、6550万年前に小惑星だと思われる巨大天体の衝突によって発生した。
この時の大量絶滅では、地球上の恐竜と、すべての生物種の大半が消滅した。「最も簡単な解釈は、生命は常に進化しているということです。進化の過程の中では、いつでも、その時点で生きている動物が比較的最近出現したものであるという可能性が高いのです」
この論文は、人類進化学の専門誌「ヒューマン・エボリューション(Journal of Human Evolution)」に掲載された。
この見解では、ある種が持続するのは一定の期間でしかなく、その後、新しいものに進化しなければ絶滅するということになる。
(後略)