免疫って何?(11)~交配分裂も殖産分化も免疫進化も、大共通項は膜タンパク質の組み換え~
前回の「免疫って何?(10)~リンパ球の起源~」では、リンパ球が何故作られたのかを記事にしました。リンパ球は脊椎動物に固有の免疫細胞ですが、有髄神経を作り出す為に生まれたシュワン細胞の登場がその背景にあります。シュワン細胞は、神経の電気伝導性を高める=跳躍伝導を実現する為に神経細胞を絶縁するための細胞です。
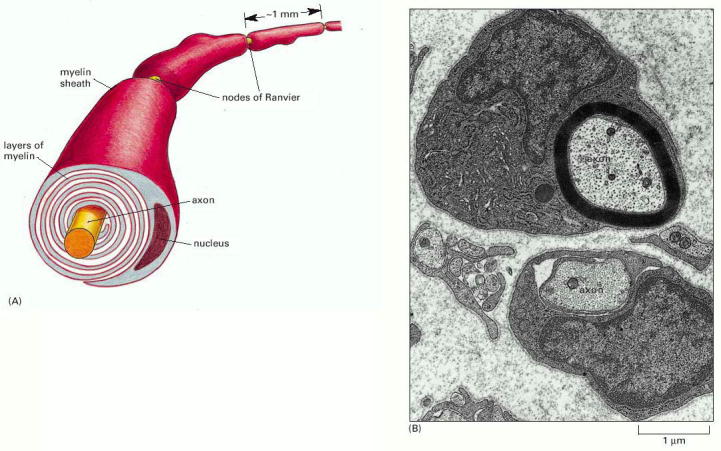
上記のように神経細胞の周りを脂質層でグルグル巻きにし体液との絶縁性を高め跳躍伝導を実現しています。このグルグル巻きの構造を作り出すためにシュワン細胞は反撥膜タンパク質を剥離させ、自己細胞の融合性を高めています。この反撥膜タンパク質を剥離させる剥離物質が他の体細胞の反撥膜タンパク質へも影響を及ぼし、ウィルスを惹き付けることとなったのが、リンパ球を必要とした理由と考えられます。
そこで今回は、もう一歩突っ込んで、そもそも膜タンパク質は単細胞からの進化の過程でどの様に組み替えられてきたのか、また脊椎動物になって、どのように膜タンパク質が変異したのかを記事(仮説)にしたいと思います。
■n体接合(ボルボックスの例)⇒同類接着タンパク質の登場
多細胞生物前夜の多細胞体制をとっているのがボルボックスでいわゆる群体と言われている生物。ボルボックスはn体(一倍体)で存在しており、鞭毛で泳ぐ体細胞と、無性生殖(まれに有性生殖)を行なう生殖細胞と役割が分化した細胞が互いにくっつきあって群体を形成している生物です。

体細胞にとってもこの現象は重要だ。胚の段階では、体細胞の鞭毛は内側に向かって生えているが、インバージョンによって外側に向く。外に向かって伸びた鞭毛で水をかいて泳げるようになるわけだ。体細胞は接着性のタンパク質を分泌し、隣り合った細胞間を埋めることで成長し、やがて親の球を破って水中に飛び出す。生命史ジャーナル2003年冬号より
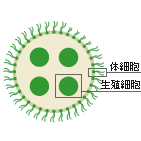
この時に細胞間の接合を果たしているのが接着タンパク質。この接着タンパク質は同じ”同類”のボルボックス同士を接着させるタンパク質なので、この群体化の実現に登場したタンパク質を同類接着タンパク質とここでは言います。
■n体融合(クラミドモナスの例)⇒同類融合膜タンパク質の登場
n体融合は群体のあたりで登場します。環境が飢餓状態に陥ると、細胞同士が一つに合体し休眠状態にはいります。その例としてクラミドモナスがあげられます。
クラミドモナスは群体を作らないボルボックス目の真核単細胞ですが、一つの膜に複数のクラミドモナスが囲まれた群体に近いものから、二つのクラミドモナスが膜融合→核融合まで進むものまであり、受精や多細胞化の原初形態といえるかもしれません。
>クラミドモナスは、染色体を一セットしか持っておらず、われわれで言えば、精子や卵と同じ。これが合体すると染色体を二セット持つわれわれの体の細胞と同じになる。つまり、飢餓状態に置かれたクラミドモナスが合体して染色体を二セット持つ細胞になったのは、ちょうど受精したのと同じことになるわけである。<
クラミドモナスは二匹合体すると厳しい環境下で休眠し、(DNAのシャッフル後)、環境条件が好転すると減数分裂してまた活発に動き回るようです。一倍体が活性状態、二倍体が休眠(防衛)状態ということでしょうか。飢餓状態で合体する、飢餓状態で休眠するというのは、(雌雄に特化・分化をとげていない)生物界を通じてかなり普遍的な適応戦略だったのではないでしょうか。るいネットより
このn体融合を引き起こすタンパク質が融合タンパク質。同類のクラミドモナスとの融合を果たすのでこれを同類融合膜タンパク質と言います。
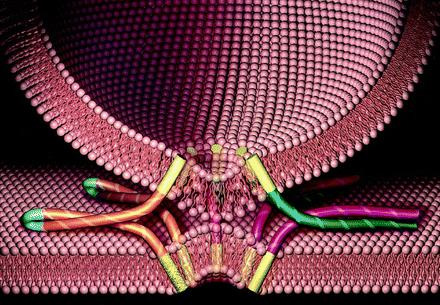
膜融合とはそもそも互いの膜同士が1.5nm以下になると自然に行なわれます。通常であれば膜表面の水分子等で1.5nm以下になることはありませんが、それを1.5nm以下に近づける為に、同類接着膜タンパク質を膜内に組み込み、互いの接着膜タンパク質同士をくっつける事で実現しているのだと思われます。つまりn体融合を実現する同類融合膜タンパク質とは、n体接合の時に獲得した同類接着膜タンパク質を膜内に組み込んだものだと考えられます。下図はそのモデル図です。
■2n融合の阻止 ⇒同相反撥膜タンパク質の登場
しかし、2n体として多細胞化するとさらに細胞間の関係に新たな課題が出てきます。n体は同類融合膜タンパク質で融合しても良いですが、2n体が同様に融合してしまうと4n体へなってしまいます。もちろん生物(植物)には4n、8n、16nと色んな生物がいますが、交配分裂による変異能の獲得によって進化してきた生物は2n体の維持が必須です。
故に2n体の細胞同士が融合しないように阻止するタンパク質が必要で、それを反撥膜タンパク質といいます。同じ同類でも同じ2n体という”相”にだけ反撥する膜タンパク質ということで、同相反撥膜タンパク質とここではいいます。この同相反撥膜タンパク質は、n体融合の時に獲得した同類融合膜タンパク質に、反撥膜タンパク質を取り付けることで融合を阻止していると考えられます。
■2n対の接合 ⇒同位同相接合膜タンパク質の登場
もう一つ多細胞化が推し進められることで出てきた課題もあります。
n体接合は先に述べた同類接着膜タンパク質によって接合されています。
本格的に多細胞化が進み、体細胞の役割分化が推し進められた各々の体細胞は、同じ体細胞同士がくっつきあう必要があります。しかし、役割の違う体細胞同士がくっつく訳にはいきません。故に、役割分化に応じた分化接着膜タンパク質が必要になります。
同じ役割を担う細胞を”同位”ということにしますが、この分化接着膜タンパク質に、上記の同類融合膜タンパク質+同相反撥膜タンパク質が組み込まれたのが、同位同相接合膜タンパク質で、これにより役割の同じ細胞とは接合し、役割の違う細胞とは同相反撥膜タンパク質によって離れる事ができているのではないかと考えられます。
■脊椎動物化 ⇒同相反撥膜タンパク質の剥離
多細胞化以降の段階で細胞間の関係に大きな変化が現れたのが脊椎動物です。脊椎動物は神経細胞、筋芽(筋肉)細胞、破骨細胞などの細胞群が登場してきます。これらの細胞の共通点は、いずれも細胞の融合が行なわれている点です。
神経系の進化は神経細胞をとりまくシュワン細胞によって跳躍伝導を実現したことは既に前回でも記述しました。シュワン細胞は自己細胞を一部融合させることで絶縁性を実現しています。また、神経系の発達と連関して筋肉の細胞である筋芽細胞も自己融合しています。
おそらく、筋細胞が運動能力を高度化させるために進化してきたものであることは間違いないであろう。そして、その進化は神経系統の進化と対を成すものであろう。筋細胞の合胞体化⇒細胞融合は、運動のための指令をより正確かつ効率的に筋細胞に伝達し、筋細胞同士が一つの統合された組織として運動するために形成されたシステムなのではないだろうか?やはり、運動ニューロンから伝えられる指令は、バラバラの細胞に個別に伝達されるより、一つの細胞に統合された合胞体に伝達した方がスムーズであろう。るいネットより
また、神経や筋肉の進化と密接に連関した破骨細胞も自己融合細胞である。
骨に存在する破骨細胞を顕微鏡でのぞいてみると,核をたくさん持った大きな細胞,つまり多核の巨細胞であることが分かります(図4)。破骨細胞がこのように多くの核を持ち,巨大化するのは細胞分裂によるものではなく,細胞融合によるものであることが示されています。その証拠の一つとして,破骨細胞の核の数は必ずしも偶数ではなく,しばしば奇数になっています。破骨細胞は単核でも骨を吸収することができるのですが,骨を活発に吸収している破骨細胞はほとんど多核の巨細胞です。なぜ多核となる必要があるのか,また融合に関与するメカニズムも不明です。ノバルティス・ファーマより
これらの細胞は2n体の融合を阻止する同相反撥膜タンパク質を剥離させ同類融合膜タンパク質とすることで、自己融合を実現していると思われます。この反撥膜タンパク質を剥離させる物質もまたタンパク質と思われますが、神経や筋肉、骨格などの自己融合=剥離タンパク質の影響によって、他の体細胞の反撥膜タンパク質も剥離しやすい状況下になり、他の体細胞との融合可能性が高まったのだと思われます。
この傍証事例にウィルス感染細胞の融合能があります。ウィルス感染細胞は、正常細胞と融合する性質を持っています。これはウィルス感染細胞とは反撥膜タンパク質が剥離された状態であることを物語っています。
ウィルスの起源は諸説色々ありますが、ウィルスとは何かと一言でいえば、遺伝子の切れ端。遺伝子の切れ端は単細胞時代から存在してますが、多細胞化さらには雄雌分化が進み遺伝子変異の増大によってミスした遺伝子の切れ端が増大する可能性も高まります。その意味ではウィルスとは自身が生み出したミスの産物と捉えるのが自然であり、大きく見れば同類の遺伝子です。
つまり反撥膜タンパク質を喪失し、同類融合膜タンパク質となった細胞というのは、ウィルスの同類融合の可能性を格段に増大させるものだったといえるかもしれません。これがリンパ球の登場を必要とした背景です。
■まとめ
今日は膜タンパク質に注目して仮説を立てました。ここで言いたかったのは、n体接合も、交配分裂も、多細胞化もウィルス誕生も、免疫細胞も、全ては膜タンパク質の改良・組み換えの産物であるという大共通項がある点です。リンパ球登場後、さらにウィルス変異に対応した抗体を作り出しますが、これもウィルスに対応した膜タンパク質の組み換えです。そういった意味で、膜タンパク質の組み換え・塗り重ねとは、人類風に言えば認識の塗り重ねと同義であるといえます。
さて、次回は最終回。観念機能を獲得した人類が、今後自然の摂理に則って正しく進化していくためには、どのような視点が必要なのか。これまでのまとめを行い、この免疫シリーズの最終回にしたいと思います。

トラックバック
このエントリーのトラックバックURL:
http://www.seibutsushi.net/blog/2010/07/991.html/trackback
いつも面白い追及ありがとうございます☆
>>何故身体が大きくなったのかは、今後の追求点の一つかもしれませんね
確かに類人猿(初期人類)から現代に至るまでの
人類の身長やその他肉体進化の理由(外圧適応)は
どうであったのかはとっても気になりますね(^ω^)
>>現代人に繋がるこの二次的成熟性はエレクトスの時に獲得したのではないかと思われます。
そもそもどのようにして発掘した化石から「二次的成熟性」
だということが分かるのでしょうか?
もし、何か分析方法等が載っていれば教えて下さいm(__)m
また他の真猿や原猿では、出産直後の脳の容量と、
成長後の脳の容量は変らない(≒二次的成熟性は人類のみ)
のでしょうか??