中心小体と同じ構造をもつ基底小体
真核生物の中心小体にはなぞが多いがその糸口になりそうなのが基底小体。中心小体と基底小体はその構成要素と構造が同じ。基底小体とは何か?それを調べてみた。
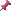 基底小体は鞭毛、繊毛の骨格を作っているチューブリンの基底部に存在
基底小体は鞭毛、繊毛の骨格を作っているチューブリンの基底部に存在
鞭毛、繊毛の骨格を作っている(チューブリンが繋がった)微小管はどんな構造かというとこれが面白い。
基底小体の話からは少しそれますが、鞭毛、繊毛の骨格である微小管は、細胞質微小管とは構造が少し違い、1対の単微小管を中心に9個の二連微小管が輪になって並んでいる。(その断面図は下図を参照)
<鞭毛の断面図>細胞生物学/南江堂より
鞭毛、繊毛の動きは波を打つような局面的な動きをしますが、この動きを
実現しているのが微小管同士をつなぐ連結タンパク質とダイニン。
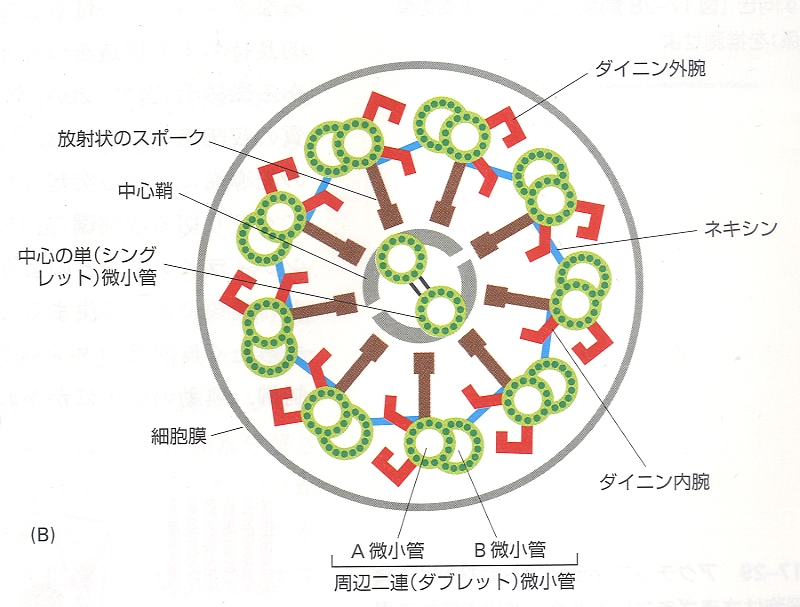
ダイニンというのは二連微小管にくっついている動くヤツ。ATPを動力源にしてダイニンの足が動くイメージです。(下図の赤色部分)連結タンパク質は二連微小管を連結しているものです。
隣り合う二連微小管がタンパク質で繋がっていない場合、ダイニンの運動によって二連微小管はお互いが滑りあう運動をします。これが連結タンパク質によって繋がれていると、ダイニンの運動により、二連微小管は屈曲した形になります。これが鞭毛、繊毛の波打つ動きを実現しています。(下図のAが滑りあう運動で、Bが波打つ運動)
すごい! ![]() よくできてるな~と関心しちゃいます。(私だけ?? 🙄 )
よくできてるな~と関心しちゃいます。(私だけ?? 🙄 )

<屈曲運動の図>細胞生物学/南江堂より
で、本題の基底小体ですが・・・
この波打つ動きをする微小管の根元に基底小体がいます。
基底小体の構造は、周辺微小管が三連微小管で構成され、中心微小管は存在しない。その形態は中心小体と全く同じで、実際に微小管形成中心として働く基底小体もあるようです。
中心小体は2個の三連微小管が90度直角になった配置となっていますが、(基底小体は色んな配置があるのか?そんな記事もあるが・・)、もう少し調べてみないと解りませんが、基底小体と鞭毛の接続のしかたはこんな感じになっています。
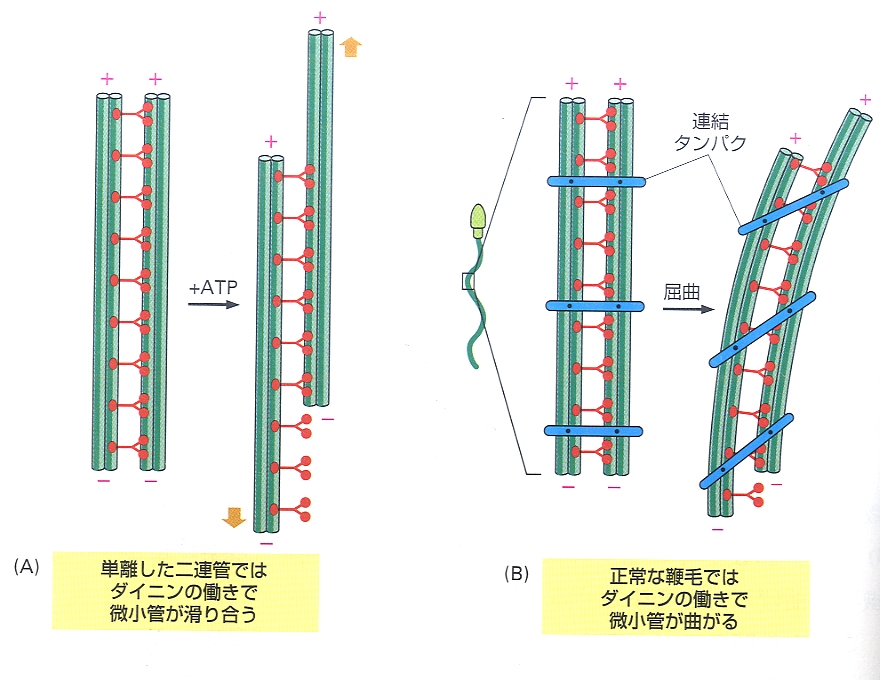
下図は黄金色藻のオクロモナス目の藻類の鞭毛装置の図。(もっと良い生物の例があればいいのですが・・・)
1、2が鞭毛で、その根元にあるのが基底小体。基底小体は鞭毛を伸長させる役割をもっている。またこの基底小体を基点にして、細胞小器官の配置が行われている。
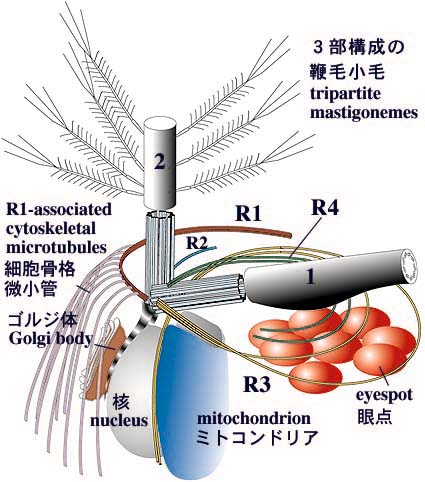
こちらのサイトから頂きました。
また、【きまぐれ生物学(雑記-分子細胞学)】にこのような記述があります。
中心小体を中心に配置を決める(2007.07.04)
細胞における細胞小器官の配置は一見乱雑でも,相対的な配置が厳密あるいは一定の範囲で決められています。 特に鞭毛の配置などは細胞運動や動物の正常な発生にも関わるために,厳密に決められる必要があります。 Feldman et al. (2007) はコナミドリムシ(Chlamydomonas reinhardtii; 緑藻植物門オオヒゲマワリ目)の変異体を用いて, 鞭毛を含め複数の細胞小器官の配置が中心小体(centriole)に依存して決まっていることを明らかにしました。
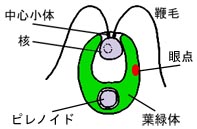
コナミドリムシは遺伝学が利用できることからモデル生物として使われていて,鞭毛を持って泳ぐ単細胞藻類です。 細胞は前端に一対の鞭毛を持ち,その直下に 1 個の核を持ちます。細胞の後半と前半の縁の部分は葉緑体が占めていて, 特に後半部(基部)には 1 個の大きなピレノイドと呼ばれる構造があります。著者らはこの藻類の中心小体に着目して研究を進めました。
コナミドリムシの細胞の模式図
中心小体は 3 本の微小管が癒合した三連微小管が 9 セット,真ん中の車輪状構造を囲むように並んだ構造で, この他にも様々な修飾を受けているようです。中心小体は通常は鞭毛の付け根にあって基底小体と呼ばれ,鞭毛を伸長させる役割を持ちます。 また分裂時には 1 対の中心小体がねじれの位置に配位して,周辺の基質と共に中心体(centrosome)と呼ばれる構造を作ります。
つまり、細胞小器官の配置を決めるのが中心小体の主な役割で、とりわけ細胞運動に関わる重要な鞭毛配置は、直接中心小体から骨格が形成されている、といったところでしょうか。
基底小体とは、単に呼び方が違うだけで、構造も役割も中心小体と同じモノ。違うのは中心小体から直接、鞭毛の骨格となる微小管を伸長させている点のようにも思えます。
ここまで調べてみて、じゃあ何で中心小体は2個直角に配置されているの?原核生物には基底小体はあるの?原核生物の中心小体原基の役割は鞭毛と関係あるの?などよくわからない事が山積みですが、今後の追及とさせていただきます。 

トラックバック
このエントリーのトラックバックURL:
http://www.seibutsushi.net/blog/2009/01/640.html/trackback
トラックバック(1)
コメント4件
iwaiy | 2009.02.07 20:50
KAMさん、こんにちは。
コメントありがとうございます!
細胞性粘菌の形態形成(←面白そうで興味あります)とGタンパク質の関係というのは知らなかったので驚きです。
Gタンパク質についてはもうちょっとつっこんで考えてみようかなと思います。特に細胞分裂との関係のあたり・・・
あとこれまでは、ヌクレオチド=RNAやDNAの材料といった一面的な認識だったのですが(それはそれであるとして)、ヌクレオチドは細胞内での様々な作用機序に不可欠の物質でもあるというのが、今回の記事を書いてのもうひとつの気づきでした。
KAM | 2009.02.09 21:42
iwaiyさんへ
生物物理を学んでいた立場からすると、非対称な細胞分裂は、「自己組織化システムの対称性の破れ」として捉えたくなります。
かつて、細胞集団の数が変わっても、細胞分化の比率が一定に保たれる仕組みを説明す数理モデルを見たことがありますが、そういったモデルでは、あるひとつのパラメータを変化させていくと、「一様な状態」が不安定化して、「2つの安定状態」へ分化していきます。
http://ci.nii.ac.jp/naid/110002237857/
細胞をいくつか取り去ると、再分化が起こるのも再現できます。
多細胞システム全体の安定性として、分化や比率制御が起こるときに、Gタンパク質が重要な役割をはたしているような気がしています。
iwaiy | 2009.02.14 21:21
KAMさん、たいへん示唆に富むコメントありがとうございます☆
知識や切り口が自在で、生物素人の私としては「なるほどな~」と思うことが多いです。
細胞の非対称分裂は、多細胞化(=
細胞の専門分化と統合)の基本ですが、確かに不思議な現象ですよね。それとヌクレオチド⇔タンパク質の関係がわかると面白いですよね!
iwaiyさん、こんにちは。
いつも興味深く読ませていただいています。
私は、10年ほど前、細胞性粘菌の形態形成を、数理モデルを使って研究していたのですが、その数理モデルの中心を担っていたのがGタンパク質でした。
そのモデルでは、三量体というのが決定的に重要で、3つの変数によって、1つの変数の時間発展が決まるところから、3次の非線形性がでてきて、1つの数理モデルで興奮性や振動状態を再現できていました。
細胞性粘菌の場合は、シグナル伝達物質であるcAMPが、細胞間の情報伝達としても使われ、
cAMPが受容体につくことで、活性化され、cAMPが合成される仕組みになっていて、正のフィードバックがかかり、それが、多細胞体が自己組織化されるカギになります。
Gタンパク質の重要性は、調べていけば、まだまだ出てきそうですね。