変異転写の仕組み?(中間整理)
ゲノム解読の進展により、どのようにして生物が外圧に適応する変異が起こるのか?の仕組みが明らかになりつつあります。
DNA変異を引き起こす変異転写(逆転写)の仕組みがあるのではないか?という問題意識のもと、これまでに分かったことを整理してみます。
<リンクより引用>
気になる続きはポチッとお願いします。

![]()

●平常時はRNA干渉によってDNA変異は制御されている
>ヒトの遺伝子は2~3万と推定される現在であるが、ここ何十年も生物学者を悩ませ続けた問題がある。それはタンパク質をコードするDNAは全DNAのたった2%程度なのである。それが大海に浮かぶ島のようにのように点々と散らばっている。あとのDNAは一体何のためにあるのだろう?学者は答えに窮してこれをがらくたDNA(Junk DNA)と呼ぶようになった。
>ここ数年の間に小さな二重鎖RNA分子がもとになって遺伝子の発現を抑制するRNA干渉(RNA Interference, RNAi)のメカニズムが次第に明らかにされるようになった。がらくたDNAはなんとこのRNAをコードしているらしい。
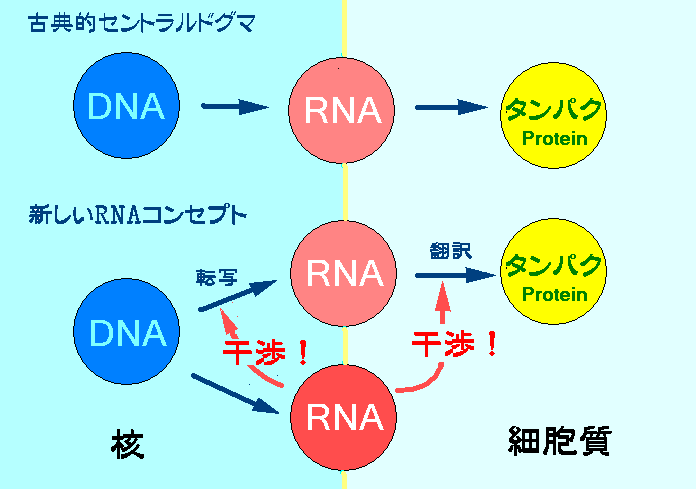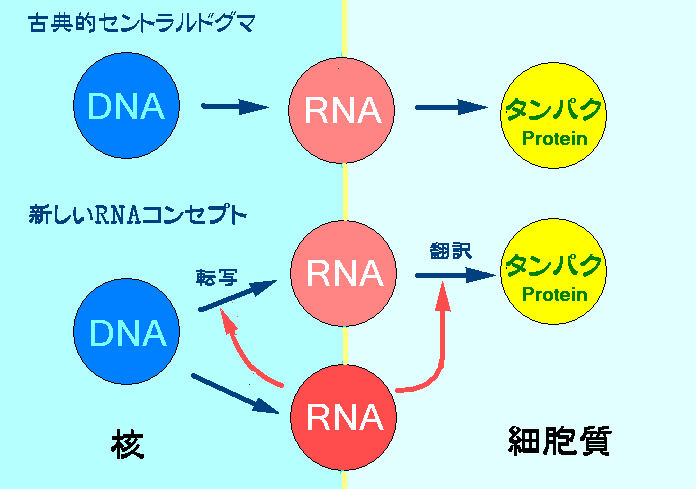
>発生、分化、成長の各段階で2万といわれる遺伝子は適時適所で順序良く発現されなければならない。実はこの遺伝子の発現をコントロールしているのがジャンクDNAからつくられる小さなRNAによるRNA干渉らしい。microRNAとかsiRNAと呼ばれる小さなRNAは細胞質で遺伝情報を運ぶメッセンジャーRNA (mRNA)を切断することによりタンパク質への翻訳を阻害する。RNAがRNAを調節しているのである。(翻訳レベル遺伝子サイレンシング)
>さらに最近に至ってこのRNAには細胞核のDNAに作用してDNA情報をRNAに写す転写レベルの阻害を示すことも次第に明らかにされて来ている。RNAがDNAを沈黙させているのである。(転写レベル遺伝子サイレンシング)
<リンクより引用>
>多細胞生物の外来性ウイルス感染防御機構として認識さえているRNA干渉現象(RNAi)だが、慶應義塾大学のグループが、ゲノムに多数存在しているトランスポゾン(ゲノムを動き回る遺伝因子)の動きを封じるために重要な働きをしていることを明らかにした。
<リンク参照>
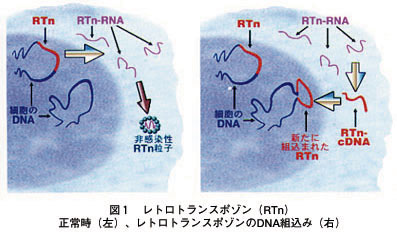
>トランスポゾンは転位性遺伝因子(動く遺伝子)の一つで、DNA 上のある部位から他の部位へ移動するDNA 単位
>トランスポゾンの転位する部位は一定ではなくランダムに転位するため、その制御は困難
>宿主染色体中に挿入されると挿入変異によりタンパク質の合成が停止したりタンパク質の変異を生じたりします
>生物において環境変化が起 こったときに脱メチル化がDNA上に起こるため、トランスポゾ ンが動き出し、変異が起こる
※メチル化:メチル基(CH3)が結合すること。酵素によって触媒される。脱メチル化はその反対でメチル基が外れること。<ウィキペディア参照>
>逆転写酵素がRNAを鋳型としてDNA鎖を合成する際、端まで合成した後に折り返してもう一方のDNA鎖も合成してしまう
>進化が必要なほど過酷な条件になると、細胞内でトランスポゾンの高い頻度の移動がみられる
<リンクより引用>
>ヒトではゲノムの3 0%以上がSINE やLINEなどのレトロポゾンである。mRNA から逆転写されて生じる「プロセシングされた偽遺伝子」が多数存在する点もヒトゲノムの著しい特徴である。
>その増幅の最盛期は各々およそ4000 万年前と推定される。このようなプロセシングされた偽遺伝子と Alu の同時期の爆発的増幅は、ある特定のLINE1 サブファミリーが細胞質RNA を逆転写する機能 を一時的に亢進させた結果ではないかと考えている。
>この時期は、原猿類から分岐した真猿類 が、新世界ザルや類人猿へと放散を遂げる直前であり、「レトロポジション爆発」が高等霊長類進化に及ぼした影響に興味が持たれる。
<リンク:「ヒトゲノム中での偽遺伝子の増幅と進化」(86/148頁)より引用>
※レトロポゾン(レトロトランスポゾン):自分自身をRNAに複写した後、逆転写酵素によってDNAに複写し返されることで転移する。(トランスポゾンがカット&ペーストであるのに対し、レトロポゾンはコピー&ペースト。)
※SINE:逆転写されたRNA 分子を表現する短いDNA 配列。ヒトゲノムの13.5% を占める。
※Alu:最も良く見られるSINE 配列。約300 塩基対の長さがあり、蛋白質コード配列を全く含まない。
※LINE(長鎖散在反復配列):逆転写されたRNA 分子を表現する長いDNA配列。偽遺伝子と呼ばれる。ヒトゲノムは約500,000 (ゲノム全体の21%) のLINE を含む。
<ウィキペディアより>
・平常時は「小さなRNA」によるRNA干渉によってトランスポゾンの動きが封じ込まれており、DNA変異は制御されている。多細胞生物では、この仕組みによって遺伝子が適時適所で順序良く発現する。(RNA干渉が解除されれば、トランスポゾンが活性化し、DNA変異が促されると考えられる。)
・外圧が高くなると、脱メチル化によってトランスポゾンの移動が活性化する。
・ヒトはレトロポゾンが多い。サルからヒトへの進化はレトロポゾンの爆発的増大によった可能性がある。
○仮説
「外圧△⇒何らかのシグナル伝達(HMG等?)⇒脱メチル化⇒(?)⇒「小さなRNA」の働き⇒RNA干渉解除⇒トランスポゾン活性化⇒変異転写(逆転写)⇒DNA変異促進」
という基本構造ではないか?
★外圧△でレトロポゾンも活性化する?(レトロポゾン→RNA→cDNA→DNAへの逆転写、表紙の図解参照)
※cDNA:相補的DNA。cDNAの塩基配列は、イントロン(タンパク質をつくるエクソンを分断している配列)を含まない。
●残課題
★RNA干渉解除の仕組みはどうなっているのか?「小さなRNA」の働き?
★RNA干渉解除の仕組みに脱メチル化が関わっているのか?
★そうであるとすれば、脱メチル化を引き起こすスイッチ(シグナル伝達)はどうなっているのか?
★ひょっとしたら、そのスイッチ役がHMGタンパク質等の変異転写を担うタンパク質である可能性はないか?
★レトロポゾン活性化の仕組み?トランスポゾンとレトロポゾンの変異転写の役割はどう違うのか?

トラックバック
このエントリーのトラックバックURL:
http://www.seibutsushi.net/blog/2008/10/592.html/trackback
コメント5件
管理者 | 2010.03.08 16:29
コメントありがとうございます。
上記の点を記事を書いた者に伝達します。
著作権者 | 2010.04.30 15:24
3/3のコメントの件です。
出典の記載をお願いします。本ブログ、他では出典の明記など適切に行われていると思います。この図の出典の明記もお願いします。
*明記後は、コメント削除して頂いて構いません。
著作権者 | 2010.07.29 15:59
中段の
“古細菌の細胞膜の特徴は、その脂質がエーテル結合である点です。というよりも、その点こそが、古細菌を他の生物と分け隔てている最大の特徴です。”という文章の上にある図の出展を明記して下さい。重ねてお願いします。
*明記後は、コメント削除して頂いて構いません。
blogger0 | 2010.07.31 19:53
>著作権者様
細胞膜構造と極性脂質の図の出典を明記しました。何度かご指摘をいただいていたにも関わらず、修正が遅くなり、申し訳ございません。
遅くなりましたが、図を引用させていただき、ありがとうございました。今後とも、よろしくお願いいたします。
細胞膜構造と極性脂質の図に出典が明記されていません。出典を記載して頂けませんか?