2008-11-05
卵割期の細胞周期
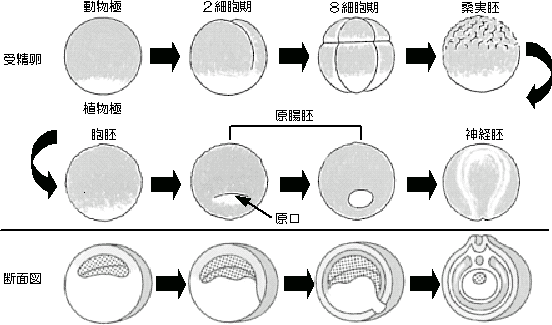
先日の記事で、多細胞生物における細胞周期は増殖因子と呼ばれる分裂を開始させる因子と、分裂を制御する細胞周期エンジン(サイクリン・CDK)によって統合されていることが分かった。
リンク
この細胞周期が原核生物でどのようになっているのか?は大変興味深く、これを調べていくことで、初期生物の細胞分裂システムに近づくことができる。
その中間形態として多細胞生物における初期細胞分裂=卵割(受精卵の細胞分裂)について調べてみました。 

![]()


リボソームRNAと核小体

今日は、RNAポリメラーゼがRNA合成を開始するために必要な、基本転写因子(タンパク質で出来ている)について調べてみました。
まず、RNA合成について簡単に復習してみましょう。
タンパク質を合成する場合、RNAポリメラーゼがDNAよりタンパク質のもととなる情報(ATGCの塩基の並び)を読み取り、その情報をもとにmRNAが出来ます(転写)。このmRNAの塩基の並びに対応した塩基を持つtRNAが集合します。tRNAは、アミノ酸を背負っています。このアミノ酸が集まり、リボゾームでタンパク質が合成されます(翻訳)。
原核生物の場合、この転写と翻訳をほぼ同時に行います。真核生物の場合、mRNAが核膜から細胞質へと移動して核外で翻訳を行います。
このRNAを作るRNAポリメラーゼへ転写の開始位置を指示しているのが、転写因子です。
紹介する記事は、リボゾームRNA合成に使われる転写因子が、原核生物~初期真核生物~植物で共通のタイプ(B型転写因子)を使っているという報告です。しかし、菌類と動物では、B型以外の転写因子に変わっているようです。
(なぜ変わったのか?非常に気になりますが、これは今後の追求課題とさせてください。)
また、この転写因子は、初期真核生物でも植物でも核内の核小体に集合しています。
おそらく、原核生物段階でも核小体のように転写因子を含むRNA合成系が集合している部分(核小体の前駆体)があったと、私は考えています。
原核生物の転写と翻訳がほぼ同時に行われるように、RNA系の合成も早いスピードで行われているはずです。核膜を持たない原核生物の場合、環状DNAのRNA系情報周辺に核小体前駆体はあるのではないでしょうか。
では以下、新たな基盤転写(RNA 合成)系の発見 原始生物シゾンで解明されたリボゾームRNA 合成系進化のミッシングリンク より、リボゾームRNAと転写因子、核小体の関係です。
いつものブログランキング応援もよろしくお願いいたします。
上図は「原核生物のリボソームの立体構造」です。画像は、こちらからお借りしました。
原核生物→真核生物でのアクチンとチューブリンの機能の逆転
原核生物(真正細菌)から真核生物に至る進化の中で、細胞骨格タンパク質であるアクチンとチューブリンの細胞内での機能が逆転しています。
具体的には、「染色体分配-細胞質分裂」の機能の逆転です。
まずは、どのように機能が逆転しているのか、逆転のしくみについて紹介します。
応援宜しくお願いします。
![]()
変異転写の仕組み?(中間整理)
ゲノム解読の進展により、どのようにして生物が外圧に適応する変異が起こるのか?の仕組みが明らかになりつつあります。
DNA変異を引き起こす変異転写(逆転写)の仕組みがあるのではないか?という問題意識のもと、これまでに分かったことを整理してみます。
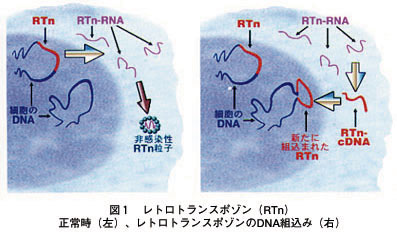
<リンクより引用>
気になる続きはポチッとお願いします。
![]()
遺伝子を制御するncRNAの協働作業
こんにちは。
今日は、ノンコーディングRNA(タンパク質に翻訳されないRNA)の役割に関する、注目の科学ニュースを紹介します 
気になる続きはポチっと押してからどうぞ
![]()
ヌクレオチドの生成と生物史

画像はコチラから
MASAMUNEです 8) 。
前回の記事からの続きです。
DNAやRNAはヌクレオチドからつくられています。
今回は原始生物はどうやって有機物(=ヌクレオチド→DNA、RNA)を獲得したか?そしてなんでヌクレオチドが遺伝情報を伝える基を担っているかをまとめた本があったので紹介します 😀 。
生物史を考えることのイミって??
こんにちは 
再び登場しました、初心者  のyukieです
のyukieです 
前回私が登場してからも、その前もず~っと 
このブログでは生物の仕組みに対する追求  (今は分子生物学的な記事が多いですね)
(今は分子生物学的な記事が多いですね)
が続いている訳なんですが
私は時々専門用語が多すぎて 
難しい~~っ 😥 てなってしまうことがあります…


(画像はこちらよりお借りしました)
そんなとき、思い出すようにしているのが、
初めて生物の仕組みを追求することに可能性を感じた瞬間!!
今日は、それをみなさんと振り返って 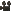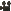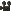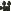 みたいと思います
みたいと思います
 応援よろしくお願いいたします
応援よろしくお願いいたします
![]()
セルトリ細胞のY染色体が精子の遺伝子を制御している
生物史ブログの「精子の乳母!?『セルトリ細胞』の役割って??」に寄せられていた、以下のコメントについて調べてみました。
>こんな記事みつけました。
↓
性分化機構の解明
哺乳類の精子発生における体細胞側でのY染色体の役割について、以下の2点を明らかにした。
1)精巣決定遺伝子Sryによる精巣への分化決定以後、思春期での半数体形成(円形精子細胞)までは、体細胞側 (精巣、下垂体、視床下部を含め体中の全ての体細胞)でY染色体は不要である。
2)円形精子細胞から精子までの精子形成に、Y染色体上のDdx3y, Uty, Ubely1, Eif2s3y, Jarid1d(全てあるいはこの一部)がセルトリ細胞で重要であり、幾つかの精子形成関連遺伝子のセルトリ細胞での発現を制御することにより精子への形態形成を支持していることが推測された。
・・・
上記だけでは判然としませんが、
セルトリ細胞におけるY染色体って、何か重要な働きをしているのかもしれませんね。
復習になりますが、セルトリ細胞は、精子の元になる精原細胞が精子に成長していくのを助けている細胞です。
この画像は京都大学のニュースリリースからお借りしました
セルトリ細胞のY染色体がどんな役割を果たしているのか、興味のある方は応援お願いします。
![]()
酵素の反応調節
こんにちわアリンコです。この前頂いたコメントにも応えなきゃと思いつつ、本日は、ちょっとおいとかせていただいて、地道に酵素の活性調節の仕組みについて投稿させていただきます。
真核生物の酵素調節には大きく2つの方法が存在します。
1:酵素タンパク質が他の生体分子と可逆的に作用することによる酵素活性の変化
2:酵素タンパク質が修飾されることによる酵素活性の変化
1や2の調節の例として、フィードバック阻害が挙げられます。
一般に触媒反応の反応速度は、基質濃度と生成物濃度により影響を受けるのですが、酵素反応の場合、ある複数の段階からなる代謝経路において、酵素の直接の基質、もしくは生成物以外の代謝生成物が酵素の反応速度を制御する場面があります。
特に、代謝生成物が過剰になったときに、生成物が何段階か上流過程の酵素反応を阻害することで産生を抑制する調節過程の事をフィードバック阻害と呼んでいます。
フィードバック制御で代表的なものとしては、アロステリック効果と共有結合修飾による調節が挙げられます。
アロステリック効果って??共有結合修飾って?と思った方はポチっと押して続きをご覧下さい。
![]()
RNAポリメラーゼのはたらきを制御する「σ因子」
RNAを合成する役割を担っている、RNAポリメラーゼという酵素があります。
この酵素のはたらきを調べている中で見つけた、RNAポリメラーゼのはたらきを制御している「σ因子」について紹介します。
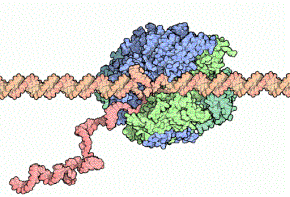
RNAポリメラーゼが、遺伝子を読み取りながらmRNAを合成するようす
😀 ![]() 😀
😀