NationalGeographic [1]より写真引用
・皆(生物学者も含んだ現代人=近代の情報知識を学生時分に身に付けた人々)は、このような疑問を持った事は無いだろうか?「単細胞生物から多細胞生物へと進化・移行出来たのは何故か?」と。そして、「そこを繋ぐ要因は何なのであろうか?タンパク質?分裂?結合?吸収?…」と。
・しかし、単細胞生物と多細胞生物の境界線をどう線引きするか?といった疑問は、そもそもの立脚点が「まず多細胞生物という地位ありきの発想」になっているのではないかと思うのです。
・これって、人間という自我による驕り以外の何者でも無いのかもしれません。
・つまり、近代に確立(観念的に固定する為に強引にでも決められた)された一定の知識・思考で物事を考えてもなかなか真実は見えてこない(∵固定的な○○ありきという発想では全くの未明なものの解明に対しては無力である)。
・だから、「事実のみを立脚点とした素人的な発想こそ」が真実を追究するに当って、最も求められている要素なのでは無いだろうかと最近至る所で感じています。
・さて、今回も事実のみを立脚点とした素人的な発想を目標にして、冒頭・タイトルでも少し述べている「立襟べん毛虫(単細胞生物)から襟細胞(多細胞生物であるカイメンの一細胞)への進化過程」を追求・展開していきたいと思います。
↓応援クリックお願いします。
 [2]
[2]
![]() [3]
[3]
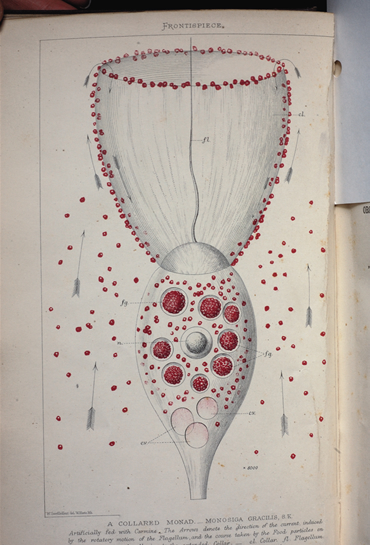
○立襟べん毛虫の位置付~たくさんある単細胞生物の中でもなぜ立襟べん毛虫を選ぶのか?~
立襟鞭毛虫は、多細胞生物(後生生物)に最も近い単細胞生物として注目されている。多細胞生物の起源については、従来から①群体起源説(鞭毛虫類の集合・群体化)と②繊毛虫類起源説(単細胞の多核化を経て多細胞化したとする考え)の2説があったが、近年の分子系統仮説からは、①群体起源説が有力と考えられている。
立襟鞭毛虫はカイメンの襟細胞によく似ていることから、古くから両者の近縁関係が指摘されていたが、最近になって分子系統学的に裏付けられた。
遺伝子解析の結果、従来、多細胞動物特有だと思われていた遺伝子が立襟鞭毛虫に多数発見されている。
われわれは、多細胞生物には多細胞独特の遺伝子があって、そうした遺伝子が多細胞らしさを形作っていると信じてきたが、その考えはどうやら捨てなければならないようだ。非常に多数の遺伝子重複が立襟鞭毛虫と動物の分岐以前に起きていたらしい。
立襟鞭毛虫がつくるタンパク質にも、多細胞動物の体にしかないと思われていた数種類のタンパク分子が見つかった。その中には細胞間のコミュニケーションを担うものもあった。これらのタンパク質の一部は通常、細胞同士がでたらめにくっつくのを防いでいるが、立襟鞭毛虫は、餌となる細菌をつかまえるためにこの接着タンパク質を使っているのではないかと考えている研究もある。
るいネット:立襟鞭毛虫とカドヘリン(多細胞生物の原点?) [4]
より引用。
○カイメンの細胞(襟細胞)の特徴
カイメンの内層:胃層
・襟細胞:水流を作る、捕食、精子を作る。
生殖方法は有性生殖と無性生殖があり、有性生殖の場合は襟細胞が精子を作り、原生細胞が卵子となる。無性生殖の場合は原生細胞が胞子のような役割を果たし、出芽により別の個体がつくられる。
カイメンの細胞は個体識別まではしておらず、同種、異種だけを見分けている。
最も原始的な多細胞生物であるカイメンは、細胞どうしが互いを特異的に認識しあって結合したものであることが、20世紀初めにH. V. Wilsonにより見つけられた。その後、実に90年経た20世紀末になって、カイメンは細胞どうしが糖鎖と糖鎖、糖鎖とタンパク質の相互作用によって結合し個体を作り上げていることが明らかとなった。
その中でも主要にカイメンの細胞結合を担っていると考えられているのがカドヘリンというタンパク質によるカドヘリン結合。同じ型のカドヘリンどうしは結合することができるため、同じ型のカドヘリンを持つ細胞どうしが接着する。
るいネット:カイメンの認識機能 [5]
より引用。
○「バイオフィルム」と「微生物とそのコロニーの変遷」
この節では、微生物とそのコロニーの詳細を説明する。
1.細菌付着
2.EPSを分泌しはじめる
3.バイオフィルムが形成される
4.バイオフィルムは厚みを増し、コロニーが巨大化する
5.内部が過密になると、コロニーが破壊され、細菌が放出される
細菌が付着と脱離を繰り返しながら、徐々にバイオフィルムが形成される。バイオフィルムのコロニーには、複数種の微生物が生息し、動的平衡を保つ。棲む微生物は、環境により異なるが、細菌類以外の微生物が生息している場合も多い。単一種のみで形成されるコロニーは、自然界には稀である。形成後のバイオフィルムも、常に脱離や溶菌が起こっているため、安定したものではない。バイオフィルムという呼び名は、極相林のような変遷の終着点というより、形成された後に変化する形態すべてを指している。ある程度大きくなると、コロニーが崩壊し、細菌が放出される。
コロニー内に、細菌が高密度に生息しているため、生活型はコロニー外と異なる。EPSや構造の隙間を利用して物質のやり取りを行う、と考えられている。EPSは、構造の支持体や防護膜として機能するだけではなく、養分の運搬・保持、酵素の伝達など、内部環境の恒常性を保つ役割も担う。また、バイオフィルム内に暮らす細菌は、EPSを通じて情報伝達物質のやり取りを行うと考えられている。こういった作用により、バイオフィルム内の環境は外部と大きく異なる。そのため、バイオフィルムの内部では、細菌の形態が相変異により異なる場合がある。細菌の構成種にも違いがみられる。たとえば、バイオフィルム内の奥部で嫌気性細菌が活動している場合がある。
ウィキペディア(Wikipedia)』より、バイオフィルム [6]
より引用。
——————————————————————
最後に、「立襟べん毛虫から襟細胞への進化過程」を仮説を踏まえて整理すると、
まず立襟べん毛虫の群れは、自然外圧に応じて、どんどんそのコロニーを拡大巨大化させた結果、当然飽和点において崩壊し、小コロニー化に分裂していったものと思われる(おそらくそれを繰り返した)。
一方で、より外圧が高い場所で生きるなければ成らなかった単細胞生物の群れ=小コロニーは適応する為に、より群れる事に特化していき、糖鎖と糖鎖、糖鎖とタンパク質による接着・結合を獲得したのだろう。
そして、単細胞の群れ=コロニーが接着タンパク質によって安定的にコロニーを巨大化させる事が可能になって以降、それらの生物は多細胞化(単細胞の拡大コロニー化)したと呼ばれるように進化したのでは無いかと思う。
⇒重要なのは、全ての生物[単細胞であっても(注)そもそも単語の意味がおかしいが・・・]は存在時点で群れる生物を出発点にしており、多細胞化とは、そのコロニーを自然的な飽和点で分解・拡散しないように接着拡大していった単細胞生物群である。という事である。