今日は受精の基本について調べてみました 
ヒトの精子は、マウスやハムスター等、他の種の卵子と結合できるのでしょうか? ![]()
子供に質問されて、うっ!え~と~・・・て、なりそうな質問ですね。
結合できれば、とんでもない種ができそうだし・・・・??? 🙄
種の保存・維持とも矛盾するし・・・・??? 🙁
答えは、結合しない。
でも、なんで結合できないのでしょう?どんな仕組で?
では、ポチポチポチっと応援して、進みましょう! 
 [1]
[1] ![]() [2]
[2]  [3]
[3]
精子と卵子が受精する時に、お互いを同じ種だと認識する機能を先体反応と呼びます。
有性生殖は、オスの精子とメスの卵子を結合することにより、親とは少し違う同類他者を作り出し、外圧に適応し種を維持します。しかし、初期の多くの生物は海の中で、体外受精を行います。また、ホヤのように雄雌同体で、精子も卵子も同時に出す生物もいます。
体外受精を行う種にとって、精子が同種の卵子と結合しないと種は維持できません。ホヤのように雌雄同体の種の場合は、自分とは少し違う他者の精子または卵子と結合しないと、同類他者はつくれません。
そこで生み出した機能が、精子と卵子の膜の認識機能をもとにした、先体反応です。ヒトデ等の初期棘皮動物と私達哺乳類では、多少の仕組みは違いますが、基本的にこの膜の認識機能を使っています。
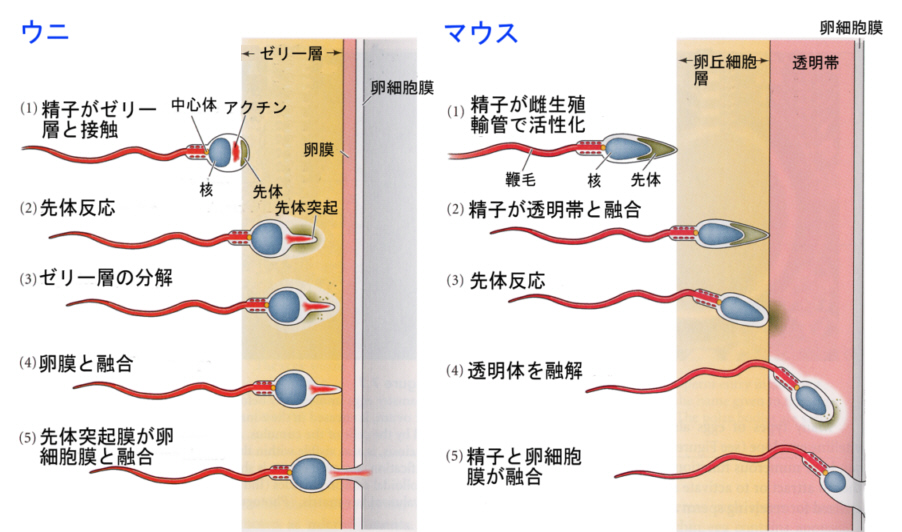
(この図は、「個体の発生と分化Ⅰ- 配偶子形成と受精 [4]」からお借りしました。
●種の認識について
上記は、棘皮動物のウニと哺乳類のマウスの精子と卵子の結合の比較です。基本的に精子と卵子の膜の認識機能を使っているのは同じです。
同種の精子と卵が認識しあって、細胞膜が融合し、精子の頭部に納められていた核と中心体が卵のなかに送り込まれます。
ウニの場合は先体反応によって先体突起が飛び出しますが、哺乳類の場合は先体反応によって酵素を放出し、これが透明帯を溶かして精子が卵細胞膜へ到達するのを助けています。
この詳細について、ウニと同じ棘皮動物の研究が分かりやすかったので紹介します。「ヒトデにおける受精メカニズムの解明」 [5]より
有性生殖は同種の異なる個体由来の遺伝子を混ぜあわせ子孫に多様性を生み出すという非常に優れた生殖戦略ですが、体外で受精を行う多くの生物にとっては異種間で生殖が起こってしまう危険性をはらんでいます。従って卵と精子が受精を成立させるためには種によって厳密に決められた幾つもの段階を経なければならないと考えられています。先体反応(Acrosome Reaction)は受精の初期段階で最も重要な種特異的認識の過程です。卵近傍に到達した精子は卵細胞を取り巻く卵黄膜(哺乳類では、透明帯)などの卵外被に種特異的に結合しその構造を認識すると、頭部先端に存在する分泌胞(先体胞)から加水分解酵素などを分泌します。これを先体反応と呼んでいて、この結果精子は卵外被を通り抜けて卵細胞膜との融合が可能になります。
ウニやヒトデのような棘皮動物の卵は、卵外被としてゼリー層と呼ばれる主に糖蛋白質から成る構造を持ち、精子はこのゼリー層と出会うと先体反応を誘起されます。更に棘皮動物の先体反応に特徴的な現象としてアクチンの重合による先体突起を形成することが知られています。
●個体の認識について
種の認識は、精子と卵子の細胞膜の認識機能を使っているのは、よく分かりました。では、個の認識はどうなっているのでしょうか?これは、一つの個体から精子も卵子も放出するホヤ等の雌雄同体の生物にヒントがありました。同じ個体から放出された精子と卵子が結合したのでは、同類他者をつくることは出来ません。
以下、「マボヤにおける自己・非自己認識反応の解明」 [6]より
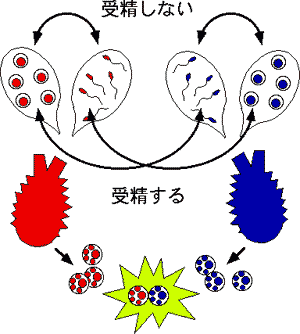
われわれ哺乳類と同じ脊索門に属するホヤ類は海に住む固着性の動物で、そのほとんどは雌雄同体です。マボヤやカタユウレイボヤでは、繁殖期になると同一個体が卵と精子の両方を同時に海水中へ放出しますが、同一個体由来の両配偶子は受精しないという現象が知られています。これを自家不和合性と呼んでいてい、この現象が有性生殖における受精の目的である「同種異個体の遺伝子を混ぜ合わせ、子孫の遺伝的多様性を生み出す」ことを保障してます。これらのホヤでは同種異個体由来の配偶子どうしならば、ほとんど全て受精することから、配偶子が同種異個体を厳密に識別できる能力をもっていると考えられています。(中略)
われわれも含めて脊椎動物は高度な細胞性免疫機構をもっていて、臓器移植における絶に見られるように同種異個体を識別することができます。進化的な位置を考えると、ホヤにおける同種異個体の識別機構が脊椎動物における細胞性免疫機構の起源となっている可能性は十分考えられます。
また、同種異個体の識別という現象はカイメンやイソギンチャクなど系統学的に離れた動物でも広く存在することから、多細胞生物に普遍的な性質とも位置づけられていて、これらが類似した分子基盤に基づく機構かどうかについても興味が持たれます。
雌雄同体であるホヤの頃に獲得した個の識別機能は、私達の免疫機構に繋がっている可能性が高いようです。
また、個の認識機能も、種を維持するためにつくられたんですね。これは、一番の気づきでした。