細胞の進化は生殖細胞と体細胞に分かれてゆきます。
生殖細胞の接合は、初めは同形配偶子という性差のない生殖細胞どうしが接合するものでした。
そのうちに、卵子と精子という異形配偶子が接合するというように進化してゆきます。
<ムチモの異形配偶子:NHK高校講座「生物」 [1]より引用>
今日は、なんで卵子と精子に分かれたのか?を考えてみます。
続きはポチッっと。
![]() [2]
[2]
 [3]
[3]
●ニハイチュウの両性生殖腺
生殖細胞が、いつごろから卵子と精子に分かれたのかははっきりしない。
しかし、ここに紹介するニハイチュウという面白い生物がいる。
ひょっとすると、このあたりの生物が、卵細胞と精細胞に分かれた原点的な生物かも知れない。
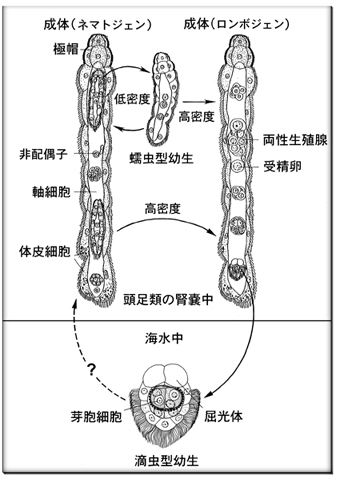
<大阪大学大学院:古屋秀隆氏のHP [4]より引用>
ニハイチュウは、主にタコやイカの腎臓に棲む多細胞生物。体長数ミリメートル。細胞数は、22個前後。最多でも50個にも及ばない。無性生殖(蠕虫型幼生)と有性生殖(滴虫型幼生)の両サイクルがあり、2種類の幼生(胚)がみられることことから二胚虫(2つの胚をもつ動物)と呼ばれる。
腎臓中での個体数が少ないときは、無性生殖(蠕虫型幼生)で個体数を増す。個体数が増加すると、有性生殖(滴虫型幼生)を行う。有性生殖では両性生殖腺が形成され、そこで自家受精した受精卵から滴虫型幼生が発生する。滴虫型幼生は尿とともに海水中に泳ぎ出て、新しい宿主に寄生すると考えられている。
両性生殖腺は、器官構造ではなく、卵系列の細胞群と精子系列の細胞群の集合体のようなもの。非配偶子から配偶子が分化する。非配偶子から3~4回の分裂で、「卵原細胞」、「精原細胞」および「両性腺の軸細胞」と呼ばれる3種類の細胞が形成される。
その後、精原細胞は軸細胞の細胞質内に取り込まれ、そのなかで精子を形成する。一方、卵原細胞は軸細胞の周囲を取り囲むように分裂し、卵を形成する。軸細胞内で成熟した精子は軸細胞内から抜け出て、外表部にある卵と出会い自家受精する。
<以上大阪大学大学院:古屋秀隆氏の研究 [5]参照>
進化の系統的には、原生動物(ミドリムシ、アメーバ、マラリア原虫、ゾウリムシなど)と後生動物(海綿動物、節足動物、脊索動物など)との中間に位置する(中生動物)とされているが、はっきりしない。(研究途上。)後生動物の海綿が、9億年前ぐらいの起源と言われているので、それより前の原始的な生物であることは間違いないだろう。
ニハイチュウの場合は、卵子に比べた精子の数は1.5倍程度であり、まだその大きさの差も大きいものではないらしい。しかし、「卵原細胞」、「精原細胞」および「両性腺の軸細胞」が見られるという点は、れっきとした異形配偶子への分化である。個体数が低密度の時は、無性生殖を行い、高密度になれば有性生殖(異形配偶子接合)を行うというのは、有性生殖(異形配偶子接合)が、生存圧力が高い時にとられる方法であることを示唆している。
●では、卵子と精子に分かれたのはなんで?
おそらく、異形配偶子の形成は、多細胞化と密接に関係している。異形配偶子による有性生殖が普遍的に見られるのは多細胞生物になってからである。多細胞生物の受精卵には、単細胞生物に比べてはるかに大量の栄養分と遺伝情報を蓄積し、なおかつ受精による遺伝子組み換えを実現するという機能が必要になる。
同形配偶子でも、大型化すれば大量の栄養分と遺伝情報を蓄積することは可能であろう。しかし、その分運動能力は劣ることになる。運動能力が劣れば、配偶子どうしが接合して遺伝子を組換えるという可能性も低下する。生殖細胞の大型化と運動能力の高度化とは相反する要請である。
そこで、卵子⇒大型化による栄養分の蓄積、精子⇒運動能力の高度化という役割分化を推し進めたのだろう。
実際、異形配偶子の場合は、進化してゆくにつれて、卵子と精子ではその大きさや運動機能の差が大きくなってゆく。動物の卵子は、大型化して栄養分をたくさん蓄積できるようになっているが、運動能力は殆どない。精子は卵子に比べて著しく小さく、栄養分を蓄える機能は殆どない。あるのは、遺伝子を蓄えた頭部と運動機能(ミトコンドリアと鞭毛等)だけである。